Unit - 4
Alkaloids
4.1.1 Natural occurrence:
Many natural compounds can be found in nature. Alkaloids appear to be unique among the several groups of naturally occurring organic compounds, including carbohydrates, lipids, proteins, amino acids, anthocyanins, flavonoids, and steroids. What distinguishes them? They're made up of amino acids and can be synthesised by plants and animals as secondary metabolites. In living organisms, these compounds play an important role. Alkaloids have long been thought to be extremely valuable to humans, despite the fact that they are secondary metabolites, which may mean that they are ineffective. In relatively small doses, alkaloids had strong biological effects on animal and human species. Alkaloids are used in a variety of foods and beverages, as well as in stimulant medications. Anti-inflammatory, anticancer, analgesics, local anaesthetic and pain reliever, neuropharmacologic, antimicrobial, antifungal, and a variety of other behaviours were demonstrated. Alkaloids are useful in medicine and other aspects of human life as diet foods, vitamins, and pharmaceuticals. Alkaloids are also essential compounds in organic synthesis for the development of new semisynthetic and synthetic compounds with potentially higher biological activity than their parent compounds.
Since alkaloids have such a wide structural diversity, there is no single method for extracting them from natural raw materials. Most approaches take advantage of the fact that most alkaloids are soluble in organic solvents but not in water, and that their salts have the opposite tendency.
Alkaloids are found in almost all plants. Their mixture is extracted first, followed by the separation of individual alkaloids. Until extraction, the plants are thoroughly ground. The majority of alkaloids are found in raw plants as organic acid salts. The alkaloids extracted can remain salts or transform into bases. The alkaloid bases are extracted using organic solvents such as 1,2-dichloroethane, chloroform, diethyl ether, or benzene after the raw material has been processed with alkaline solutions. Weak acids dissolve the impurities, converting alkaloid bases to salts that can be washed away with water. An aqueous solution of alkaloid salts is rendered alkaline again if possible, then treated with an organic solvent. The procedure is repeated until the desired purity level is reached.
The raw plant material is treated by a poor acidic solution in acidic extraction (e.g., acetic acid in water, ethanol, or methanol). The alkaloids are then converted to basic forms, which are extracted using organic solvents (if the extraction was performed with alcohol, it is removed first, and the remainder is dissolved in water). As previously mentioned, the solution is filtered.
Alkaloids are isolated from their mixtures using varying solvent solubility and reactivity with different reagents, or by distillation.
Researchers have reported a variety of alkaloids in insects, with solenopsins, which are alkaloids found in the venom of fire ants, receiving the most attention. Solvent immersion of live fire ants or centrifugation of live ants accompanied by silica-gel chromatography purification are both effective methods for extracting insect alkaloids. Based on their absorbance peak around 232 nanometers, tracking and dosing the extracted solenopsin ant alkaloids has been identified as possible.
4.1.2 Chemistry and Biological Activities of Flavonoids:
Introduction
Flavonoids are a wide group of polyphenolic compounds with a benzo-pyrone structure that are found in plants. The phenylpropanoid pathway is used to make them. According to research, secondary phenolic metabolites, such as flavonoids, are responsible for a wide range of pharmacological activities. Flavonoids are hydroxylated phenolic compounds that plants are known to produce in response to microbial infection. 3. Their actions are governed by a set of rules. Flavonoids' chemical properties are determined by their structural class, degree of hydroxylation, other substitutions and conjugations, and polymerization degree. 4. The potential health benefits derived from the antioxidant activities of these polyphenolic compounds have sparked renewed interest in these compounds. Flavonoids' functional hydroxyl groups scavenge free radicals and/or chelate metal ions to mediate their antioxidant effects. 5. and 6. Metal chelation may be crucial in preventing the generation of radicals that damage target biomolecules. 7, 8, 9. Flavonoids are thought to have health-promoting properties as a dietary ingredient due to their high antioxidant potential in both vivo and in vitro systems. 9, ten. Flavonoids have the ability to activate defensive enzyme systems in humans. Flavonoids have been shown to protect against a variety of infectious (bacterial and viral diseases) and degenerative diseases such as cardiovascular disease, cancer, and other age-related diseases in a number of studies 2, 9, 10. In this study, the processes involved in flavonoids' defence are discussed separately. Flavonoids also serve as a secondary antioxidant protection mechanism in plant tissues that have been subjected to various abiotic and biotic stresses. Flavonoids are found in the nuclei of mesophyll cells as well as ROS production centres. They also control plant growth factors including auxin 11. Biosynthetic genes have been assembled in a variety of bacteria and fungi to boost flavonoid production 12. The structural aspects of flavonoids and their protective functions against a variety of human diseases are discussed in this study. Flavonoids' functions in plants, as well as their microbial production, have been identified.
Chemistry of Flavonoids
Flavonoids are a class of natural compounds found in plants with varying phenolic structures. Oranges were used to isolate a new drug in 1930. Vitamin P was given to it because it was thought to be a part of a new class of vitamins at the time. It was later discovered that this material was a flavonoid (rutin), and over 4000 different flavonoids have been known to date 13.
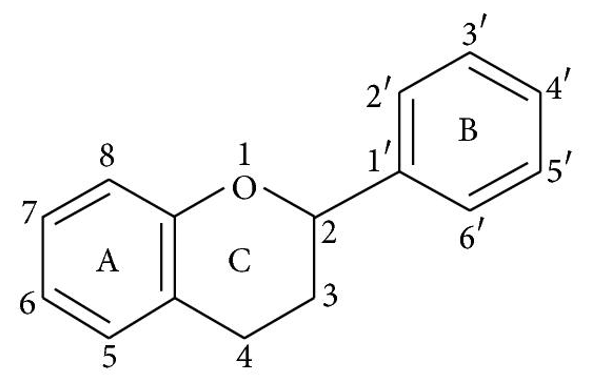
Flavonoids have a fifteen-carbon skeleton, which is made up of two benzene rings (A and B in Figure 1) connected by a heterocyclic pyrane ring (C). Flavones (e.g., flavone, apigenin, and luteolin), flavonols (e.g., quercetin, kaempferol, myricetin, and fisetin), flavanones (e.g., flavanone, hesperetin, and naringenin), and others are only a few examples. Table 1 depicts their general systems. Individual flavonoids within a class differ in their degree of oxidation and substitution pattern of the C ring, whereas individual compounds within a class differ in their substitution pattern of the A and B rings 13.
Table. Structure of flavonoids.
Flavonoids come in a variety of forms, including aglycones, glycosides, and methylated derivatives. The aglycone structure is the most fundamental flavonoid the framework (Figure 1). A -pyrone (flavonols and flavanones) or its
dihydroderivative is a condensed six-member ring with the benzene ring (flavonols and flavanones). Flavonoids (2-position) and isoflavonoids (2-position) are divided by the position of the benzenoid substituent (3-position). Flavonols differ from flavanones in that they have a 3-position hydroxyl group and a C2–C3 double bond 40. The positions 3, 5, 7, 2, 3′, 4′, and 5′ are often hydroxylated in flavonoids. The alcohol group's methyl ethers and acetyl esters are known to exist in nature. The glycosidic linkage is usually found in positions 3 or 7 when glycosides are formed, and the carbohydrate may be L-rhamnose, D-glucose, glucorhamnose, galactose, or arabinose 41.
Spectral Characteristics of Flavonoids
Spectroscopy studies on flavonoids have shown that the majority of flavones and flavonols have two big absorption bands: The B ring absorption is represented by Band I (320–385 nm), while the A ring absorption is represented by Band II (250–285 nm). Functional groups attached to the flavonoid skeleton can cause absorption to change from 367 nm in kaempferol (3,5,7,4′-hydroxyl groups) to 371 nm in quercetin (3,5,7,3′,4′-hydroxyl groups) and 374 nm in myricetin (3,5,7,3′,4′,5′-hydroxyl groups) 42. Flavones are distinguished from flavonols by the absence of a 3-hydroxyl group. According to their UV spectral characteristics, flavanones have a saturated heterocyclic C ring with no conjugation between the A and B rings. 42. Flavanones have a high Band II absorption maximum between 270 and 295 nm, respectively 288 nm (naringenin) and 285 nm (taxifolin), with just a shoulder at 326 and 327 nm for Band I. When a monosubstituted B ring is present, Band II appears as a single peak (270 nm), but when a di-, tri-, or o-substituted B ring is present, it appears as two peaks or one peak (258 nm) with a shoulder (272 nm). Since anthocyanins have a distinct Band I peak in the 450–560 nm region due to the hydroxyl cinnamoyl system of the B ring and a Band II peak in the 240–280 nm region due to the benzoyl system of the A ring, their colour varies with the number and location of the hydroxyl groups 44.
Flavonoid Rich Food and Medicinal Plants
Flavonoids are the most common and widely distributed group of plant phenolic compounds, with flavonoids found in virtually all plant parts, particularly photosynthesising plant cells. They are a big component of flowering plants' colouring. Flavonoids are a necessary component of both human and animal diets. Table 2 lists several food sources that contain various flavonoids groups. Flavonoids cannot be synthesised by humans or animals because they are phytochemicals 45. As a result, flavonoids present in animals are derived from plants rather than being biosynthesized in the wild. The most common flavonoids in foods are flavonols. Flavonoids in food are responsible for colour, flavour, fat oxidation prevention, and vitamin and enzyme safety 46. Soy isoflavones, flavonols, and flavones are the flavonoids present in the largest concentrations in the human diet. While catechins are found in most fruits and some legumes, their concentrations range from 4.5 to 610 mg/kg 47. Depending on the methods used, food preparation and processing may reduce flavonoid levels. Orange juices, for example, were found to contain 81–200 mg/L soluble flavanones in a recent study, while cloud content was 206–644 mg/L, implying that flavanones are concentrated in the cloud during processing and storage 48. Because of the large variety of available flavonoids and their widespread distribution in different plants, as well as the complex ingestion of flavonoids in humans, accurate estimate of the average dietary intake of flavonoids is difficult 49.
There has recently been a wave of interest in medicinal plants' therapeutic potential, which may be due to their phenolic compounds, specifically flavonoids 50, 51. Humans have been consuming flavonoids for around 4 million years, since the beginning of human life on the planet. They have a wide range of biological properties that benefit human health and help to minimise disease risk. During atherosclerosis, oxidative alteration of LDL cholesterol is thought to play a key role. Glabridin, a major polyphenolic compound contained in the Glycyrrhiza glabra (Fabaceae) plant, inhibits LDL oxidation via a process involving free radical scavenging. 51. Several epidemiological studies have suggested that drinking green or black tea may lower blood cholesterol levels and blood pressure, thereby reducing the risk of cardiovascular disease. Flavonoids, as flavorants, colourants, and antioxidants, are known to affect the consistency and stability of foods. 53 and 54. Flavonoids found in berries have been shown to benefit people with Parkinson's disease and enhance memory in the elderly. In hypertensive rats 55, the total flavonoid fraction of Astragalus complanatus had an antihypertensive impact. Consumption of antioxidant flavonoids has been shown to be inversely linked to the risk of dementia 56. Table 3 lists some of the medicinal plants that are high in flavonoids.
The therapeutic effectiveness of flavonoids can be influenced by their solubility. With the exception of an occasional instance of allergy, the poor solubility of flavonoid aglycones in water, combined with their limited residence period in the intestine and lower absorption, humans do not experience acute toxic effects from flavonoid intake. The low solubility of flavonoids in water makes their medicinal applications difficult. As a result, semisynthetic, water-soluble flavonoids such as hydroxyethylrutosides and inositol-2-phosphatequercetin have been linked to the treatment of hypertension and microbleeding 57.
Metabolism of Flavonoids in Humans
The physicochemical properties of dietary flavonoids liberated from food by chewing can influence their absorption, including molecular size, configuration, lipophilicity, solubility, and pKa. The flavonoid may be absorbed directly from the small intestine or may first pass through the colon. It could be determined by the flavonoid's composition, specifically whether it is a glycoside or an aglycone. Except for the catechin subclass, most flavonoids are found in plants bound to sugars as b-glycosides (Figure 2). Flavonoid glycosides must be converted into aglycan type 58 before they can be consumed by the small intestine.
(a)
(b)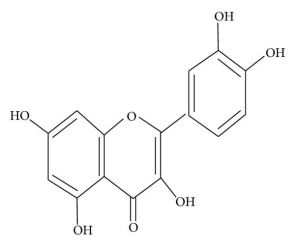
Figure: Structure of (a) flavonoid glycoside and (b) aglycone flavonoid.
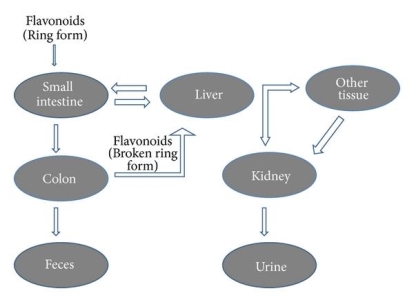
The intestinal Na+-dependent glucose cotransporter (SGLT1) 58 transports hydrophilic flavonoid glucosides like quercetin through the small intestine. Another theory is that flavonoid glucosides are hydrolyzed by lactase phloridzin hydrolase (LPH), a -glucosidase found on the outside of the small intestine's brush boundary membrane. The liberated aglycone can then be absorbed by the small intestine 59. This LPH enzyme's substrate specificity differs greatly across a wide variety of flavonoids 60 glycosides (glucosides, galactosides, arabinosides, xylosides, and rhamnosides). The glycosides that aren't substrates for these enzymes are transported to the colon, where bacteria can hydrolyze flavonoid glycosides while simultaneously degrading the liberated flavonoid aglycones 61. Since the colon's absorption potential is far lower than that of the small intestine, only a minor amount of these glycosides can be absorbed.
After absorption, flavonoids are glucuronidated, sulfated, or methylated in the liver, or metabolised to smaller phenolic compounds 62. Except for catechins 63, no free flavonoid aglycones can be contained in plasma or urine as a result of these conjugation reactions. The bioavailability of these flavonoids varies significantly depending on the food source; for example, quercetin absorption from onions is four times greater than that from apple or tea 64. Intestinal microflora degrade flavonoids secreted with bile in the intestine and those that cannot be absorbed from the small intestine in the colon, as well as the flavonoid ring structure (Figure 3). Under the influence of acidic conditions in the stomach, oligomeric flavonoids can be hydrolyzed into monomers and dimers. Larger molecules make it to the intestine, where bacteria degrade them. Flavonoid glycosides' bioavailability is largely determined by their sugar moiety. Bioavailability has been shown to be reduced by dimerization. Isoflavones have the maximum bioavailability of all the flavonoids subclasses 65. The flavonoid content of green tea is rapidly absorbed, as shown by their elevated levels in plasma and urine. Soon after ingestion, they penetrate the systemic circulation and induce a substantial rise in plasma antioxidant status 66.
Biological Activities of Flavonoids
Antioxidant Activity
Flavonoids have a variety of biochemical properties, but the ability to act as antioxidants is the property that almost any group of flavonoids is best known for. The arrangement of functional groups around the nuclear structure determines flavonoids' antioxidant activity. Several mechanisms of antioxidant activity, such as radical scavenging and metal ion chelation, are heavily influenced by the configuration, substitution, and total number of hydroxyl groups. 67, 4, Since it donates hydrogen and an electron to hydroxyl, peroxyl, and peroxynitrite radicals, stabilising them and giving rise to a relatively stable flavonoids radical 68, the B ring hydroxyl configuration is the most important determinant of scavenging of ROS and RNS.
(1) suppression of ROS formation (either by inhibiting enzymes or chelating trace elements involved in free radical generation); (2) scavenging ROS; and (3) upregulation or preservation of antioxidant defences are some of the mechanisms of antioxidant action. 69 and 70. The majority of the mechanisms described above are involved in flavonoid action. Any of the results mediated by them may be the product of a combination of radical scavenging activity and enzyme functions interacting. Microsomal monooxygenase, glutathione S-transferase, mitochondrial succinoxidase, NADH oxidase, and other ROS-producing enzymes are all inhibited by flavonoids 71.
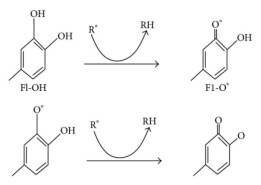
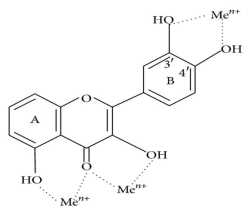
Oxidative stress causes lipid peroxidation, which is a common side effect. Flavonoids protect lipids from oxidative damage in a variety of ways. 51, 5, By reducing hydrogen peroxide and generating the highly reactive hydroxyl radical, free metal ions boost ROS production. Flavonoids (Fl-OH) can reduce strongly oxidising free radicals (redox potentials in the range 2.13–1.0 V) such as superoxide, peroxyl, alkoxyl, and hydroxyl radicals by hydrogen atom donation due to their lower redox potentials (Figure 4(a)). Flavonoids inhibit free radical production due to their ability to chelate metal ions (iron, copper, etc.) 70, 72. Iron-chelating and iron-stabilizing properties of quercetin are well-known. Trace metals bind to unique locations in flavonoid structures' rings 73. Figure 4 depicts the binding points (b).
(a)
(b)
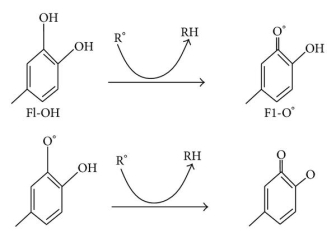
Figure: (a) ROS scavenging by flavonoids (Fl-OH) and (b) trace metal binding sites, where indicates metal ions.
A 3′,4′-catechol structure in the B ring significantly improves lipid peroxidation inhibition. Flavonoids are the most powerful scavengers of peroxyl, superoxide, and peroxynitrite radicals because of this property. 4. In vitro, epicatechin and rutin are strong radical scavengers and lipid peroxidation inhibitors 74. When the B ring of flavonoids with catechol groups is oxidised, a relatively stable orthosemiquinone radical is formed, which is a good scavenger. On oxidation, flavones lacking the catechol system produce unstable radicals with low scavenging potential 75. Flavonoids with an unsaturated 2-3 bond in conjugation with a 4-oxo function are more potent antioxidants, according to the literature, than flavonoids missing one or both features. The aromatic nucleus has a resonance effect when the A and B rings are conjugated, which gives the flavonoid radical stability. The presence of both elements, as well as other structural features, enhances flavonoids' ability to scavenge free radicals. 76th.
By allowing conjugation between the aromatic rings and the presence of a free 3-OH, the flavonoid heterocycle contributes to antioxidant activity. Coplanarity and conjugation are compromised when a 3-OH annulus is removed, compromising scavenging capacity 77. The OH groups on the B ring are thought to form hydrogen bonds with the 3-OH, allowing the B ring to align with the heterocycle and A ring. The effect of a 3-OH is strengthened by the presence of a 3′,4′-catechol due to this intramolecular hydrogen bonding, elucidating the potent antioxidant activity of flavan-3-ols and flavon-3-ols with the latter function. In general, O-methylation of flavonoids' hydroxyl groups reduces their radical scavenging potential 76.
Flavonoid (flavonoids glycosides) occurrence, position, composition, and total number of sugar moieties all play a role in antioxidant operation. Aglycones are more powerful antioxidants than their glycoside counterparts. According to some studies, the antioxidant properties of tea flavonol glycosides decreased as the amount of glycosidic moieties increased by 78. Though glycosides are typically weaker antioxidants than aglycones, a glucose moiety may often improve bioavailability. Flavonoid glycosidic moieties at the 3- or 7-position 79 are the most common in the diet. The efficacy of procyanidins against a number of radical species increases as the degree of polymerization increases. Procyanidin dimers and trimers are more effective against superoxide anion than monomeric flavonoids. Tetramers are more active than trimers against peroxynitrite and superoxide-mediated oxidation, and heptamers and hexamers have slightly better superoxide scavenging properties than trimers and tetramers 80.
Hepatoprotective Activity
Several flavonoids have been found to have hapatoprotective properties, including catechin, apigenin, quercetin, naringenin, rutin, and venoruton 81. Hepatic clinical symptoms may evolve as a result of a variety of chronic diseases, such as diabetes. In the liver of diabetic mice, glutamate-cysteine ligase catalytic subunit (Gclc) expression, glutathione, and reactive oxygen species (ROS) levels were found to be lower. Anthocyanins are gaining popularity due to their ability to protect against a variety of diseases. According to Zhu et al. 82, the anthocyanin cyanidin-3-O-glucoside (C3G) increases hepatic Gclc expression by raising cAMP levels, which cause protein kinase A (PKA), which then upregulates CREB phosphorylation, promoting CREB-DNA binding and increasing Gclc transcription. Hepatic ROS levels and proapoptotic signalling are reduced when Gclc expression is increased. C3G therapy also reduces hepatic lipid peroxidation, prevents the release of proinflammatory cytokines, and protects against hepatic steatosis 82.
Silymarin is a flavonoid derived from the seeds and fruit of the milk thistle Silybum marianum with three structural components: silibinin, silydianine, and silychristine (Compositae). Silymarin has been shown to induce DNA-dependent RNA polymerase 1 enzymatic activity and subsequent RNA and protein biosynthesis, resulting in DNA biosynthesis and cell proliferation, which leads to liver regeneration only in damaged livers 83. In normal livers, silymarin increases proliferating hepatocytes in response to FB1 (Fumonisin B1, a mycotoxin developed by Fusarium verticillioides) induced cell death while having no effect on cell proliferation. Silymarin's pharmacological properties include cell membrane permeability and integrity control, leukotriene inhibition, ROS scavenging, suppression of NF-B function, protein kinase depression, and collagen development 84. Cirrhosis, ischemic injury, and toxic hepatitis caused by various toxins such as acetaminophen and toxic mushroom 85 can all be treated with silymarin.
Flavonoids isolated from Laggera alata were found to have hepatoprotective properties against carbon tetrachloride (CCl4)-induced injury in primary cultured neonatal rat hepatocytes and in rats with hepatic harm. Flavonoids at concentrations ranging from 1 to 100 g/mL increased cell viability and prevented CCl4 86-induced hepatocyte aspartate aminotransferase (AST) and alanine aminotransferase (ALT) leakage. In an in vivo study, flavonoids at 50, 100, and 200 mg/kg oral doses significantly decreased serum levels of AST, ALT, total protein, and albumin, as well as liver levels of hydroxyproline and sialic acid. Flavonoid 86 therapy also improved the weakened liver, according to histopathological tests.
Flavonoids have been shown to be effective and secure in the treatment of hepatobiliary dysfunction and digestive symptoms such as fullness, lack of appetite, nausea, and abdominal pain in several clinical studies. In HepG2 cells, flavonoids from Equisetum arvense, as well as hirustrin and avicularin isolated from other sources, have been shown to protect against chemically induced hepatotoxicity 87, 88.
Antibacterial Activity
Flavonoids are known to be produced by plants in response to microbial infection, so it's no surprise that they've been found to be effective antimicrobial substances in vitro against a wide range of bacteria. Antibacterial activity has been identified in flavonoid-rich plant extracts from various species 70, 72, 89, 90. Apigenin, galangin, flavone and flavonol glycosides, isoflavones, flavanones, and chalcones are some of the flavonoids that have been shown to have antibacterial efficacy. No. 91.
Antibacterial flavonoids may have several cellular targets rather than a single action site. Nonspecific forces such as hydrogen bonding and hydrophobic effects, as well as covalent bond formation, are used by them to form complexes with proteins. As a result, their antimicrobial mode of action may be linked to their ability to inactivate microbial adhesins, enzymes, cell envelope transport proteins, and other proteins. Microbial membranes can also be disrupted by lipophilic flavonoids. 92 and 93.
Due to their antimicrobial activity, catechins, the most reduced form of the C3 unit in flavonoid compounds, have been extensively researched. In vitro antibacterial activity against Vibrio cholerae, Streptococcus mutans, Shigella, and other bacteria has been identified for these compounds 94, 95. Catechins have been shown to inhibit isolated bacterial glucosyltransferases in S. mutans and inactivate cholera toxin in Vibrio cholera, most likely due to complexing activities 94, 96. In Proteus vulgaris, robinetin, myricetin, and ()-epigallocatechin all inhibit DNA synthesis. According to Mori et al. 97, the flavonoids' B ring can intercalate or form hydrogen bonds with the stacking of nucleic acid bases, inhibiting DNA and RNA synthesis in bacteria. In another study, quercetin, apigenin, and 3,6,7,3′,4′-pentahydroxyflavone were found to inhibit Escherichia coli DNA gyrase 98.
Antibacterial activity of naringenin and sophoraflavanone G against methicillin-resistant Staphylococcus aureus (MRSA) and streptococci. This effect can be due to a change in membrane fluidity in hydrophilic and hydrophobic regions, implying that these flavonoids can reduce the fluidity of membranes' outer and inner layers 99. The connection between antibacterial activity and membrane interference lends credence to the theory that flavonoids exert antibacterial activity by reducing bacterial cell membrane fluidity. In the flavanone structure, the 5,7-dihydroxylation of the A ring and the 2′,4′- or 2′,6′-dihydroxylation of the B ring are critical for anti-MRSA activity 100. Flavanones and flavones have a hydroxyl group at position 5 that is essential for their anti-MRSA activity. The antistaphylococcal activity of flavonoids belonging to the flavan-3-ol class 101 can be enhanced by substituting C8 and C10 chains. 5-hydroxyflavanones and 5-hydroxyisoflavanones with one, two, or three additional hydroxyl groups at the 7, 2′, and 4′ positions inhibited S. mutans and Streptococcus sobrinus 102 formation, according to Osawa et al.
Haraguchi and colleagues 100 investigated the antibacterial activity of two flavonoids isolated from the roots of Glycyrrhiza inflata, licochalcones A and C, against S. aureus and Micrococcus luteus. They discovered that licochalcone A prevented radioactive precursors from being incorporated into macromolecules (DNA, RNA, and protein). Because energy is needed for active uptake of various metabolites as well as biosynthesis of macromolecules, this behaviour was similar to the mode of action of antibiotics inhibiting the respiratory chain. Further research revealed that the flavonoids' inhibition site in the bacterial respiratory electron transport chain 100 was between CoQ and cytochrome. Phytoconstituents obtained from edible and medicinal plants have been shown to be effective antibacterial agents in numerous studies 103–105.
Anti-Inflammatory Activity
Inflammation is a natural biological reaction to tissue damage, microbial pathogen infection, and chemical irritants. Inflammation is triggered by immune cells migrating from blood vessels and the release of mediators at the site of injury. This is accompanied by the mobilisation of inflammatory cells, the release of reactive oxygen species (ROS), reactive nitrogen species (RNS), and proinflammatory cytokines to destroy foreign pathogens, and the repair of damaged tissues. Inflammation is normally fast and self-limiting, but abnormal resolution and persistent inflammation can lead to a variety of chronic diseases 106.
Diet, pharmacologic agents, environmental toxins, and naturally occurring food chemicals can all affect the immune system. Certain flavonoids have a direct impact on immune system and inflammatory cell activity 107. Flavonoids including hesperidin, apigenin, luteolin, and quercetin have been shown to have anti-inflammatory and analgesic properties. Flavonoids may affect specifically the function of enzyme systems critically involved in the generation of inflammatory processes, especially tyrosine and serine-threonine protein kinases 108, 109. The competitive binding of flavonoids with ATP at catalytic sites on the enzymes causes kinase inhibition. Signal transduction and cell activation processes involving immune system cells are mediated by these enzymes. Flavonoids have been shown to inhibit the expression of inducible nitric oxide synthase, cyclooxygenase, and lipooxygenase, which are responsible for the synthesis of a large amount of nitric oxide, prostanoids, leukotrienes, and other inflammatory mediators including cytokines, chemokines, and adhesion molecules 110. Phosphodiesterases involved in cell activation are also inhibited by flavonoids. The biosynthesis of protein cytokines that mediate the adhesion of circulating leukocytes to injury sites accounts for a large part of flavonoid's anti-inflammatory impact. Certain flavonoids are effective inhibitors of prostaglandin formation, a class of powerful proinflammatory signalling molecules 111.
Silymarin therapy has been shown to reverse carrageenan-induced inflammatory changes. In vitro, quercetin has been found to inhibit mitogen-stimulated immunoglobulin secretion of IgG, IgM, and IgA isotypes. At concentrations of 1–10 mM, some flavonoids have been shown to significantly inhibit platelet adhesion, aggregation, and secretion 113. The inhibition of arachidonic acid metabolism by carbon monoxide 114 has been linked to the effect of flavonoid on platelets. Certain flavonoids, on the other hand, are potent inhibitors of cyclic AMP phosphodiesterase, which may explain their ability to inhibit platelet function in part.
Anticancer Activity
Dietary factors play a significant role in cancer prevention. Flavonoids have been identified as cancer chemopreventive agents in fruits and vegetables 72, 115. Consumption of onions and/or apples, two main sources of the flavonol quercetin, is linked to a lower risk of prostate, lung, stomach, and breast cancer. Furthermore, moderate wine drinkers seem to have a lower risk of lung, endometrium, oesophagus, stomach, and colon cancers 116. The importance of fruit and vegetable consumption in cancer prevention has been well established. It has been proposed that significantly increasing consumption of these foods may result in significant public health benefits 117.
Influences on development and hormonal activities have been suggested as pathways for flavonoids' impact on the initiation and promotion stages of carcinogenicity 118. The following are the major molecular mechanisms of action of flavonoids: (1) downregulation of mutant p53 protein, (2) cell cycle arrest, (3) tyrosine kinase inhibition, (4) heat shock protein inhibition, (5) oestrogen receptor binding capability, and (6) inhibition of Ras protein expression.
In human cancers, p53 mutations are one of the most common genetic abnormalities. The inhibition of p53 expression may cause cancer cells to enter the G2-M phase of the cell cycle. Flavonoids have been shown to reduce the expression of mutant p53 protein in human breast cancer cell lines to nearly undetectable levels. 119th. Tyrosine kinases are a family of proteins that play a role in the transmission of growth factor signals to the nucleus. They are found in or near the cell membrane. Their expression is thought to play a role in oncogenesis by allowing them to bypass normal growth regulation. Drugs that block tyrosine kinase activity could be used as antitumor agents without the cytotoxic side effects associated with traditional chemotherapy. In a human phase I trial 120, quercetin was the first tyrosine kinase inhibitor studied. Heat shock proteins form a complex with mutant p53, allowing tumour cells to avoid normal cell cycle arrest mechanisms. Heat shock proteins also help cancer cells survive under a variety of physical stresses. In many malignant cell lines, including breast cancer, leukaemia, and colon cancer, flavonoids have been shown to inhibit the development of heat shock proteins 119.
The flavanol epigallocatechin-3-gallate has recently been shown to inhibit fatty acid synthase (FAS) activity and lipogenesis in prostate cancer cells, an effect that has been linked to growth arrest and cell death 116, 121. FAS expression is significantly increased in different human tumours, in comparison to most common tissues. Early in the growth of a tumour, FAS is upregulated, and this is amplified in more advanced tumours 122.
In proliferating lymphoid cells, quercetin has been shown to cause cell cycle arrest. In addition to its anticancer properties, quercetin inhibited the development of many malignant tumour cell lines in vitro. P-388 leukaemia cells, gastric cancer cells (HGC-27, NUGC-2, NKN-7, and MKN-28), colon cancer cells (COLON 320 DM), human breast cancer cells, human squamous and gliosarcoma cells, and ovarian cancer cells 119 were among the cells studied. According to Markaverich et al. 123, quercetin's inhibition of tumour cell growth may be due to its association with nuclear type II oestrogen binding sites (EBS). Quercetin, when used as an antiproliferative agent, has been shown to significantly reduce increased signal transduction in human breast cancer cells 124.
Barnes 125 has done a thorough study of genistein's anticancer effects in both in vitro and in vivo models. Genistein was found to suppress the growth of chemically induced mammary cancer without reproductive or endocrinological toxicities in a study to assess the effects of the isoflavones genistein, daidzein, and biochanin A on mammary carcinogenesis. In rats, neonatal administration of genistein (a flavonoid) was found to protect against the development of induced mammary cancer 126. Hesperidin, a flavanone glycoside, has been shown to protect rats from developing colon and mammary cancers after exposure to azoxymethanol 127. Carroll et al. 128 looked at the anticancer effects of flavonoids found in citrus fruits. Antimutagenic activity has been identified for several flavonols, flavones, flavanones, and the isoflavone biochanin A 129. It was discovered that a carbonyl function at C-4 of the flavone nucleus is needed for their activity. Antitumor effects have also been shown for flavone-8-acetic acid 130. BP-7, 8-diol-9, and 10-epoxide-2 tumorigenicity were shown to be inhibited by ellagic acid, robinetin, quercetin, and myricetin in previous studies on mouse skin 131.
A higher intake of phytoestrogens, such as isoflavones and other flavonoids, has been linked to a lower risk of prostate cancer 132. It is well established that oxidative stress may lead to cancer initiation, so potent antioxidants have the ability to slow down the development of carcinogenesis. The ability of an antioxidant to serve as an oxygen radical inactivator and inhibitor determines its anticancer potential 70, 72, 133. As a result, diets high in radical scavengers will reduce the cancer-causing effects of certain radicals 134.
Antiviral Activity
Because of their availability and potential low side effects, natural compounds are a valuable source for the discovery and production of novel antiviral drugs. Antiviral activity of naturally occurring flavonoids has been recognised since the 1940s, and there are several studies on the antiviral activity of various flavonoids. The need of the hour is to find an effective drug to combat the human immunodeficiency virus (HIV). The majority of research on antiviral compounds focuses on inhibiting different enzymes involved in the virus's life cycle. Flavonoids have been shown to have a structure-function relationship with their enzyme inhibitory activity. Flavan-3-o1 was found to be more effective than flavones and flavonones in inhibiting HIV-1, HIV-2, and other immunodeficiency virus infections by Gerdin and Srensso 135. HIV-1 infection and replication was inhibited by baicalin, a flavonoid isolated from Scutellaria baicalensis (Lamieaceae). Baicalein, as well as other flavonoids including robustaflavone and hinokiflavone, has been shown to block HIV-1 reverse transcriptase 136. Another research found that the flavone O-glycoside 137 inhibits HIV-1 entry into cells expressing CD4 and chemokine coreceptors and antagonises HIV-1 reverse transcriptase. Catechins have also been shown to suppress HIV-1 DNA polymerases. Flavonoids that inhibit HIV-1 proteinase 136 include demethylated gardenin A and robinetin. The flavonoids chrysin, acacetin, and apigenin have also been shown to prevent HIV-1 activation through a novel mechanism that most likely involves inhibition of viral transcription 138.
Synergism has been shown in various combinations of flavones and flavonols. Against the herpes simplex virus, kaempferol and luteolin have a synergistic impact (HSV). Flavonoids and other antiviral agents have also been shown to work together. Quercetin has been shown to enhance the effects of acyclovir and 5-ethyl-2-dioxyuridine against HSV and pseudorabies infection 136. Flavonols are more active than flavones against herpes simplex virus type 1, according to studies, with galangin, kaempferol, and quercetin 136 being the most active.
Zandi et al. 139 investigated quercetin, hesperetin, naringin, and daidzein's antidengue virus properties at various stages of DENV-2 (dengue virus type-2) infection and replication. In Vero cells, Quercetin was found to be the most potent against DENV-2. Several flavonoids, including dihydroquercetin, dihydrofisetin, leucocyanidin, pelargonidin chloride, and catechin, have antiviral activity against HSV, respiratory syncytial virus, poliovirus, and Sindbis virus 135. Antiviral mechanisms of action have been suggested, including inhibition of viral polymerase and binding of viral nucleic acid or capsid proteins 139. Table 4 shows a list of flavonoids and their effectiveness against viruses.
Role of Flavonoids in Plants
Plants have long been known to use flavonoids for a variety of purposes 140. Various abiotic and biotic factors contribute to the development of reactive oxygen species (ROS) in plants, resulting in oxidative stress. Plants' flavonoid biosynthesis is almost entirely boosted by oxidative stress. They have the ability to absorb the most energetic solar wavelengths (UV-B and UV-A), inhibit ROS development, and quench ROS once they have developed 141. When early plants transferred from the water to the land, flavonoids performed primary UV-B screening functions. The nature of substitution on different rings of flavonoids determines the extent of antioxidant potential and ability to absorb UV-wavelengths. Flavonoids with a dihydroxy B ring substituted have a higher antioxidant potential, whereas monohydroxy B ring substituted flavonoids have a higher ability to absorb UV-wavelengths 141.
Flavonoids' most volatile hydroxyl groups (7-OH in flavones and 3-OH in flavonols) are typically glycosylated. Glycosylation increases solubility in aqueous cellular environments, protects reactive hydroxyl groups from autooxidation 142, and enables flavonoids to be transported from the endoplasmic reticulum to different cellular compartments and secreted to the plasma membrane and cell wall 143. Antioxidant flavonoids have recently been discovered in the nucleus of mesophyll cells and inside ROS-generating centres, such as the chloroplast. H2O2, hydroxyl radicals, and singlet oxygen 141, 144 can all be easily quenched here.
Conditions that restrict CO2 diffusion to carboxylation sites and carboxylation efficiency can aggravate oxidative stress caused by an excess of excitation energy in the chloroplast 141, 145. Drought/salinity, low/high temperature, and nutrient scarcity are some of the environmental constraints to CO2 assimilation rate. The function of ROS detoxifying enzymes in the chloroplast 146, 147 may be significantly reduced under these conditions, which upregulates the biosynthesis of ROS scavenging flavonoids. Flavonoids' reducing properties are extremely important in plants that are under a lot of stress. These functional functions are linked to the presence of dihydroxy B ring substituted flavonoids 148 in high concentrations. Flavonoids have been proposed as a secondary antioxidant protection mechanism in plant tissues that have been exposed to various stresses. Number 141. Oxidative stress causes lipid peroxidation, which compromises the cell membrane's integrity. At the water-lipid interface, quercetin 3-O-rutinoside (rutin) can interact with the polar head of phospholipids, increasing membrane rigidity and thus protecting membranes from oxidative damage 149.
2. As Growth Regulator
Flavonoids play an important role in plant-environment interactions. Auxin movement and catabolism can be controlled by flavonoids (in the nanomolar range). Flavonoids' ability to generate auxin gradients results in phenotypes with various morphoanatomical characteristics 150. Controlling auxin movement with flavonoids may be extremely useful in stress-induced morphogenic responses in plants, such as the flight strategy of sessile organisms in unfavourable environments 151. When comparing dihydroxy flavonoids-rich species to monohydroxy flavonoids-rich species, phenotypes with remarkably different morphological traits emerge 152. In sunny environments, dwarf bushy phenotypes with few, thin, and dense leaves to direct sunlight irradiance are common, shielding leaves deep in the canopy from light-induced cellular homeostasis perturbations. Shaded plants, on the other hand, have long internodes and broad leaf lamina with reduced leaf thickness 151, and are rich in kaempferol and/or apigenin derivatives (with negligible concentrations of quercetin derivatives).
PIN (pin formed) and MDR (multidrug resistance) glycoproteins, which are involved in the cell-to-cell movement of auxin, are effectively inhibited by flavonoids at the plasma membrane. The presence of the catechol group in the B ring of the flavonoid skeleton determines flavonoids' ability to inhibit the function of the efflux facilitator PIN and MDR proteins. Furthermore, flavonoids control the activity of IAA-oxidase in a variety of ways, depending on their chemical structure. Number 153. Recent evidence of flavonoids (as well as flavonoid biosynthesis enzymes) being found in the nucleus suggests that flavonoids can modulate the function of proteins involved in cell development. As a result, flavonoids can function as transcriptional regulators 154, 155.
Microbial Production of Flavonoids
In response to the low efficiency of plant and chemical synthesis, researchers have turned to microorganisms to produce flavonoids using metabolic engineering and synthetic biology 156. The chemical synthesis of flavonoids necessitates harsh reaction conditions and the use of hazardous chemicals 157. Combinatorial biosynthesis provides an advantage for the processing of unusual and costly natural products due to rapid advancements in molecular biology techniques and the influx of genome knowledge from a variety of species. It can be used in both simple and complex transformations without the time-consuming blocking and deblocking steps that organic synthesis needs 158. Flavonoids have been developed using a variety of prokaryotes and eukaryotes, including E. coli, Saccharomyces cerevisiae, Streptomyces venezuelae, and Phellinus igniarius, a medicinal mushroom 12.
The precursor for a large number of flavonoids formed by the phenylpropanoid synthetic pathway in plants is naringenin chalcone. Fermentative synthesis of flavanones from the amino acid precursors phenylalanine and tyrosine 159 in E. coli carrying an artificially assembled phenylpropanoid pathway was the first example of a nearly complete biosynthetic pathway in plants being formed in a heterologous microorganism. The action of phenylalanine ammonia-lyase deaminates phenylalanine to yield cinnamic acid, which is the first step in the phenylpropanoid pathway in plants (PAL). Cinnamate-4-hydroxylase (C4H) converts cinnamic acid to p-coumaric acid, which is then activated to p-coumararoyl-CoA by 4-coumarate: CoA ligase. Chalcone synthase (CHS) catalyses the condensation of three acetate units from malonyl-CoA with p-coumaroyl-CoA to produce naringenin chalcone in a stepwise fashion. In vitro 160, chalcone isomerase (CHI) or nonenzymatically converts naringenin chalcone to naringenin.
2. Enhancement of Flavonoid Production
The primary molecular biology technology procedures used in the heterologous development of flavonoids are the combination of promoter and target genes, knockout of related genes, overexpression of malonyl-CoA, and construction of artificial P450 enzymes. Every phenylpropanoid pathway gene is cloned in the host under the guidance of the promoter, which frequently plays a key role in the heterologous expression of secondary metabolites. T7, ermE*, and GAL1 promoter 12 have all been used to increase flavonoid development in response to the needs of particular hosts. One of the disadvantages in the microbiological processing of flavonoids was the exceptionally low concentration of malonyl-CoA in the microbial cell. The intracellular malonyl-CoA pool was amplified by coordinated overexpression of acetyl-CoA carboxylase genes from Photorhabdus luminescens, resulting in increased flavonol output 161. In the biosynthesis of flavonoids, UDP-glucose is also an important effector. It was shown in a study in which the udg gene, which codes for UDP-glucose dehydrogenase, was knocked out. This resulted in the removal of the endogenous UDP-glucose intake pathway, resulting in an increase in intracellular UDP-glucose concentration and, as a result, an increase in flavanone and anthocyanin output 162.
The difficulty in expressing active, membrane-bound cinnamate-4-hydroxylase 163 is one of the barriers to producing flavonoids and related compounds in microorganisms by assembling biosynthetic genes to shape an artificial pathway. Because of its instability and the absence of its cognate cytochrome P450 reductase in the host, this enzyme is not efficiently expressed in bacteria. Flavonoid formation in yeasts or fungi has the advantage of being able to express functionally active microsomal cytochrome P450 enzymes, which are difficult to express in an active form in bacteria. In the flavonoid biosynthesis pathway 164, several microsomal cytochrome P450 enzymes are involved. Researchers were able to produce a larger library of natural and unnatural products by combining bacterial and eukaryotic cells in a single pot than any of the previously described systems. For the first time, de novo development of the main flavonoid intermediate naringenin from glucose was demonstrated using an engineered S. cerevisiae strain, resulting in fourfold higher concentrations than previously recorded in de novo biosynthesis studies 165, 166.
Conclusion
The use of phytochemicals, especially flavonoids, in the prevention and treatment of diseases is well known. Flavonoids are found naturally in fruits and vegetables. The physical, chemical, and physiological properties of various flavonoids found in nature vary. Flavonoids' structure-function relationship epitomises major biological activities. Many flavonoids have been shown to have antibacterial, hepatoprotective, anti-inflammatory, anticancer, and antiviral properties. In developed countries, these substances are more widely used. Biochemical methods must be used to confirm the therapeutic application of new substances. Flavonoids can now be produced in large quantities thanks to genetic modifications. More accomplishments would contribute to newer discoveries and, without a doubt, a new era of flavonoid-based pharmaceutical agents for the treatment of many infectious and degenerative diseases.
4.1.3 Assessment of Antioxidant Capacity and Putative Healthy Effects of Natural Plant Products Using Soybean Lipoxygenase-Based Methods.
Introduction
Free radical scavengers, singlet oxygen quenchers, inactivators of peroxides and other reactive oxygen species (ROS), metal ion chelators, quenchers of secondary oxidation products, inhibitors of pro-oxidative enzymes 2, and compounds that induce upregulation of antioxidative enzymes are examples of antioxidants. Natural antioxidants have received increased attention in recent decades, especially in the biological, medical, and nutritional fields, due to their putative protective roles against the deleterious oxidative-induced reactions implicated in food deterioration and the pathogenesis of many human diseases, such as atherosclerosis, diabetes mellitus, and chronic infancy. Natural and potent antioxidants are also in high demand as food preservatives and nutraceuticals/pharmaceuticals. Antioxidant Capacity (AC) assessment methods must be accurate in order to effectively search for naturally occurring antioxidants and design novel antioxidant compounds. 2.
For this purpose, a considerable number of chemical assays, coupled with highly sensitive, quick and usually automated detection technologies, have been developed (Table 1) 1,7,8,9,10,11,12,13,14,15,16,17,18,19. These assays claim to provide a quick, easy, convenient, and reliable in vitro AC determination by providing different information about the ROS–sample interaction. They typically use a chemical system that includes an oxidant (free radicals or other ROS), an oxidizable probe (not needed for certain assays), and the antioxidants being studied.
Table: List of the widespread assays to evaluate Antioxidant Capacity.
Assay | Main Mechanism | Oxidant | Probe | Detection | Ref. |
ORAC | HAT | ROO∙ | FL | Fluorescence | 1,7 |
DPPH | SET | DPPH∙ | DPPH∙ | Absorbance | 8,9 |
FRAP | SET | Fe3+ | Fe(TPTZ)22+ | Absorbance | 10 |
TEAC | SET | ABTS∙+ | ABTS+ | Absorbance | 11,12 |
HORAC | HAT | HO∙ | FL | Fluorescence | 13 |
TRAP | HAT | ROO∙ | β-PE | Fluorescence | 14,15 |
CUPRAC | SET | Cu2+ | Neocuproine | Absorbance | 16 |
Total Phenolic Assay | SET | FCR | FCR | Absorbance | 17 |
Crocin Bleaching | HAT | ROO∙ | Crocin | Absorbance | 18 |
Chemiluminescence | HAT | H2O2 | Luminol | Fluorescence | 19 |
ABTS+: radical cation 2,2′-azino-bis (3-ethylbenzothiazoline-6- sulfonic acid); CUPRAC: Cupric Ion Reducing Antioxidant Capacity; DPPH: 2,2-Diphenyl-1-picrylhydrazyl; DPPH radical; FCR: Folin Ciocalteu Reagent; Fe(TPTZ)22+: 2,4,6-Tris(2-pyr FL: Fluorescein; -PE: phycoerythrin; -PE: phycoerythrin; -PE: phycoerythrin; -PE: phycoery FRAP stands for Ferric Ion Reducing Antioxidant Power; HORAC stands for Hydroxyl (HO) Radicals Averting Capacity; ORAC stands for Oxygen Radical Absorbance Capacity; ROO stands for peroxylradical produced by 2,2′-Azobis(2-methylpropionamidine) dihydrochloride; TEAC stands for Trolox Equivalent Antioxidant Capacity; TRAP stands for Total Radical-tra
Antioxidant activities can be measured as inhibition of ROS-mediated oxidation of the probe or as equivalents of a reference antioxidant like Trolox, ascorbic acid, or other compounds. 2. Assays based on hydrogen atom transfer (HAT) reactions and assays based on single electron transfer (SET) reactions can be loosely divided into two categories, depending on the reactions involved (see also Table 1). The HAT reaction, which is most applicable to human biology 1, is characterised by a coordinated movement of a proton and an electron in a single kinetic stage, in which a free radical removes one hydrogen atom from an antioxidant and the antioxidant becomes a radical. The antioxidant eliminates the free radical by a single electron transfer in SET mechanisms. HAT and SET reactions usually occur at the same time, and the reaction mechanism is defined by the antioxidant's structure and solubility, the partition coefficient, and the solvent polarity 20.
In fact, these methods have some technical flaws, most of which are related to the chemistry used in the various assays. In fact, most of these approaches can only assess antioxidants' scavenging ability against particular types of radicals, some of which are physiologically and biologically insignificant, leaving other significant antioxidant effects unaffected. As a result, a more valid technique for a more biologically applicable AC evaluation might be a proper combination of AC assays with different chemical bases.
Nonetheless, in vitro AC determinations should be viewed with caution due to the difficulties of accurately representing the in vivo situation 2,7,21,22. To obtain reliable information on the potential of dietary antioxidants as health-promoting agents, in vitro chemical methods may not be sufficient. In particular, important parameters involved in biological settings, such as lipophilicity and bioavailability, in vivo stability, antioxidant retention by tissues, and reactivity in situ, are not taken into account in these chemical assays. Finally, there is mounting evidence that metabolic pathways linked to bioactive compounds' prevention or amelioration of chronic diseases are often dependent on enzyme/protein and/or gene expression control rather than a true antioxidant effect 23,24. AC assays have been expanded from food model systems to biological samples, cell lines, and even live tissues 2 for these purposes.
4.1.4 Methods Based on the Use of the Soybean Lipoxygenase-1 Isoenzyme: Lipoxygenase/4-Nitroso-N, N-Dimethylaniline (LOX/RNO) and
Lipoxygenase–Fluorescein (LOX-FL) Assays
Two advanced assays focused on the use of the soybean lipoxygenase (LOX)-1 isoenzyme as a device to produce different physiological radicals and to highlight different antioxidant mechanisms have been developed in order to develop novel methodologies/approaches capable of providing AC values as closely as possible resembling the in vivo response. They were given the names LOX/RNO 25 and LOX–FL 26 because they are based on the reactions of LOX-1 isoenzyme with 4-nitroso-N, N-dimethylaniline (RNO) 27 and LOX-1 isoenzyme with fluorescein (FL). The first LOX/RNO approach was used to investigate the antioxidant properties of a variety of natural products, including food-grade antioxidants, cereal and pseudocereal grains, grain-derived products, and fruits (28,29,30,31,32). The LOX–FL assay was recently established 26 by combining the advantages of the LOX/RNO process, which stem from the use of soybean LOX-1 isoform, with the high sensitivity of the ORAC assay, which stems from the use of FL as a probe 33. In comparison to the LOX/RNO process, this has the added benefit of being a technique that can be used for both in vitro measurements of food extracts and ex vivo analysis of human blood samples. As a result, it has primarily been used to calculate ex vivo AC in human blood after both short-term 34 and long-term 35 dietary antioxidant intakes.
The data obtained from in vitro and ex vivo measurements using LOX-1-based assays is provided in an up-to-date summary.
Aerobic and Anaerobic Reactions Catalyzed by Soybean Lipoxygenase (LOX)-1 Isoform and Involvement of LOX-1-Mediated Reactions in RNO Bleaching and FL Quenching
Lipoxygenases (also known as linoleate: oxygen oxidoreductase) are a wide family of non-heme, non-sulfur iron-containing fatty acid dioxygenases found in plants and mammals 36,37,38,39. These enzymes catalyse the incorporation of molecular oxygen into polyunsaturated fatty acids containing at least one 1,4-cis,cis pentadiene moiety (e.g., linoleic, linolenic, and arachidonic acids) in an area- and stereo-specific manner to generate the corresponding hydroperoxy derivatives 40. LOXs have a variety of physiological functions 41,42,43. Fatty acid hydroperoxides produced by plant LOXs can be metabolised into volatile aldehydes and jasmonates, which are important signal molecules in wound healing and defence 44; hydroperoxides are precursors of lipoxins and leukotrienes, which are involved in inflammation, asthma, and heart disease in mammals. 45 and 46.
In Scheme 1, the four consecutive dioxygenation reactions of linoleic acid (LH) to 13-hydroperoxy-linoleate (LOOH) catalysed by the soybean LOX-1 isoenzyme are shown. The non-heme iron atom (Fe) plays a key role in LOX-1 catalysis, cycling from the oxidised form (III) to the reduced form (II) 47. The first step involves stereo-selective hydrogen abstraction followed by reduction of non-heme ferric (III) iron to ferrous (II) form, which results in proton release and generation of the linoleate alkylic (L) radical within the LOX-1–iron (II) complex's substrate-binding pocket. Molecular dioxygen insertion can occur after alkylic radical rearrangement, resulting in the peroxyl radical (LOO)–enzyme–iron (II) complex. By transferring one electron from the ferrous (II) iron atom that is reoxidized to its ferric (III) form, the peroxyl radical is reduced to the corresponding anion (LOO–). The peroxyl anion is then protonated to hydroperoxide (LOOH) and released from the enzyme–iron (III) complex, allowing a new cycle to begin. The alkylic (L) radical reacts with LOOH formed during the aerobic cycle to form LH and LOO 48 when the main aerobic cycle consumes oxygen in the reaction mixture. Via Russell's mechanism 49, the peroxyl radical will produce hydroxyl radical (OH), as well as carbonyl compounds (dienals and oxodienes) 48 and singlet oxygen (1O2). By using a Fenton-like reaction, the enzyme–iron (II) can convert LOOH into hydroxyl anion (OH–) and linoleate alkoxyl (LO) radical 47. All of these radicals have been shown to cause plant pigment oxidation and bleaching 41. Furthermore, some of these reactive species (LO, LOO, OH, and 1O2, but only in the presence of imidazole 50) have been shown to induce RNO bleaching in a biochemical pathway linked to the formation of oxodiene 27. The capacity of these oxidant species to cause FL bleaching and quenching has also been demonstrated recently 26. Antioxidants working through different pathways can slow, inhibit, or even prevent the soybean LOX-1-catalyzed RNO bleaching (LOX/RNO reaction) and FL quenching (LOX–FL reaction), as shown in Scheme 1. (indicated by green arrows). Secondary antioxidant mechanisms include chelating or reducing activities of iron ion necessary for LOX-1 catalysis, singlet oxygen quenching, hydroperoxide decomposition, and direct inhibition of pro-oxidative LOX-1 apo-enzyme. As a result, unlike the majority of AC assays, the LOX/RNO and LOX–FL reactions can simultaneously detect primary scavenging potential against many different and biologically specific radical species, as well as other essential secondary antioxidant functions 25,26, even when oxygen supply is low (as it often is in vivo). As a result, methods based on the LOX/RNO or LOX–FL reactions may be able to provide more integrated and thorough information about the AC of foods 25,26.
It should be noted that assays involving inhibition of soybean LOX-induced fatty acid peroxidation have already been proposed 51,52,53 among methodologies for AC assessment of food antioxidants. These assays, unlike the LOX/RNO and LOX–FL reactions, focus on secondary antioxidant mechanisms, such as the ability to avoid or delay lipid peroxidation by decomposing or inactivating lipid hydroperoxides, chelating iron ions, or directly inhibiting the enzyme.
LOX enzyme that is pro-oxidative. An assay based on -carotene bleaching combined with LOX-induced linoleic acid peroxidation in soybeans has also been established 54. While the LOX/RNO and LOX–FL reactions have the advantage of being able to track reaction progress with inexpensive and highly water-soluble probes, the -carotene-linoleic acid co-oxidation assay uses the highly fat-soluble -carotene. As a probe, carotene was used. Furthermore, it should be noted that -carotene is an inhibitor of LOXs 55, which may result in misleading results. Finally, the LOX/RNO and LOX–FL reactions will reveal antioxidant scavenging mechanisms against a variety of biologically relevant oxidant species produced by soybean LOX-1 at low oxygen concentrations, whereas the inhibition of LOX-mediated -carotene bleaching is largely dependent on the scavenging of peroxyl radicals generated during the aerobic cycle of linoleate oxidation 56.
LOX/RNO and LOX–FL Reactions: Main Kinetic Properties
By continuously monitoring the decrease in signal, described by RNO absorbance at 440 nm, the LOX/RNO reaction can be easily photometrically measured. Similarly, as the FL fluorescence decreases (ex = 485, em = 515 nm), the LOX–FL reaction can be followed. The standard LOX/RNO or LOX–FL reaction (trace a) and simultaneous oxygen uptake measurement (dotted line, trace b) progressions are schematically represented in Figure 1A.
1st Figure
RNO bleaching and FL quenching kinetics catalysed by soybean LOX-1 (A) Examples of typical experimental traces include: I spectrophotometric (or fluorimetric) measurement of the LOX-1-dependent RNO bleaching (or FL quenching) reaction (trace a) and (ii) simultaneous polarographic measurement of LOX-1-catalyzed oxygen uptake (trace b) (dotted line, trace b). Also shown are the reaction rate (v) and the lag phase. (B) Michaelis-Menten plots show the rates of RNO bleaching and FL quenching, expressed as a percentage of Vmax. For the LOX/RNO and LOX–FL reactions, 100 percent Vmax corresponds to 0.27 0.10 A440min1 and 0.28 0.01 A485min1, respectively. The kilometres travelled are also mentioned. Data from Pastore et al. 25 and Soccio et al. 26 have been recalculated and expressed as mean value standard deviation (n = 3).
Due to linoleate dioxygenation, the addition of LOX-1 to a reaction mixture containing excess linoleate and a low oxygen concentration causes rapid oxygen consumption (trace b in Figure 1A). RNO bleaching or FL quenching, which is indicated by a rapid signal decrease as a function of time (trace an in Figure 1A), on the other hand, occurs only after a lag period of about 20 to 120 seconds 25,26. It is the amount of time required for primary aerobic LOX-1-mediated LH peroxidation to reduce the oxygen concentration in the reaction mixture to 20–50 M 27, triggering secondary anaerobic reactions that generate radical species capable of causing signal reduction (see also Scheme 1).
The highest slope of the experimental trace is used to measure the rate of LOX/RNO or LOX–FL reaction. The rate of the LOX/RNO reaction was found to be affected by the amount of LOX-1, linoleate concentration, pH, and temperature, as well as responsive to the LOX inhibitor n-propylgallate 27. Pastore et al. 25 and Soccio et al. 26 have demonstrated the enzymatic existence of both LOX/RNO and LOX–FL reactions, reporting a hyperbolic dependency of reaction rate on RNO or FL concentration, as predicted by the Michaelis–Menten equation (Figure 1B), as well as Lineweaver–Burk, Eadie–Hofstee, Eadie-Scatchard, and Hanes plots. The saturation dependence of reaction rate on RNO or FL concentration indicates that reactions take place in the LOX-1 substrate-binding pocket, resulting in a ternary enzyme–radical–RNO (or FL) complex. The ability of LOX/RNO and LOX–FL reactions to occur in the body of a biological macromolecule is an intriguing feature, as it allows for a more physiological approach to studying antioxidant effects than the most commonly used assays 25,26.
Inhibition of LOX/RNO and LOX–FL Reactions by Pure Antioxidant Compounds
The typical activity of the LOX/RNO or LOX–FL reaction in the presence of a pure antioxidant compound is shown in Figure 2A. The ability of antioxidants to inhibit LOX-1-based reactions by inducing a decrease in reaction rate is clearly shown, with increasing inhibition with increasing antioxidant concentration.
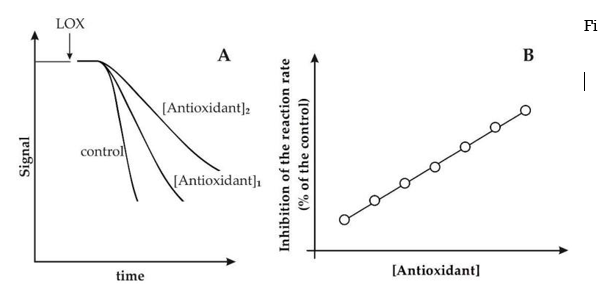
Antioxidants inhibit the LOX/RNO (or LOX–FL) reaction. (A) Experimental traces of LOX/RNO (or LOX–FL) reactions in the absence (control) and presence (two increasing concentrations) of a generic antioxidant. (B) As a function of antioxidant concentration, the inhibition is stated as a percent decrease in the LOX/RNO (or LOX–FL) reaction rate compared to the power.
However, it should be noted that some antioxidants may have a major effect on both the reaction rate and the lag phase, or may trigger a lag phase increase only 25,28. Different behaviours of different antioxidant compounds in terms of reaction rate and lag step may indicate different phytochemical antioxidant acts. Compounds that primarily inhibit the reaction rate may have a key radical scavenging function, while antioxidants' ability to exert an obvious effect on the lag process may imply an inhibition of LOX-1 hydroperoxidative activity (antiperoxidative action) 25,28. Interestingly, as shown in Figure 2B, a linear plot can be obtained by reporting antioxidant-dependent inhibition as a function of antioxidant concentration, expressed as (percent) decrease in reaction rate compared to the power. Using regular pure antioxidants, this made achieving a proper calibration curve for both LOX/RNO and LOX–FL reactions a breeze. Calibration curves for both LOX/RNO and LOX–FL reactions have been obtained using Trolox, a -tocopherol analogue with improved water solubility, as defined by the equations: respectively, (percent)inhibition = 5.838 Trolox(mM) + 11.374 (r = 0.996 ***) and (percent)inhibition = 1.920 Trolox(M) + 3.23 (r = 0.996 ***).
Several pure compounds from the main classes of antioxidants (phenolic acids, flavonoids, stilbenes, tocols, carotenoids, vitamins, etc.) were tested for their ability to affect the LOX/RNO reaction 25; additionally, the most representative endogenous antioxidants found in human serum were tested. All of the compounds tested were found to influence the LOX/RNO and/or LOX–FL assays by inducing a reaction rate inhibition with varying mechanisms and efficacy. For each antioxidant, the nature of inhibition was calculated using Ki and/or IC50 values, and the most important findings are summarised in Table.
Table: Effects of different antioxidant compounds on LOX/RNO and LOX–FL reactions.
Compound | Ki a or IC50 b | Antioxidant/Trolox c |
LOX/RNO Reaction | ||
Trolox | 7.0 ± 1.1 mM a,d | 1.00 |
Resveratrol | 1.7 ± 0.2 mM a,d | 4.57 ± 0.33 e |
Ferulic acid | 10.7 ± 2.1 mM a,d | 0.99 ± 0.11e |
Gallic acid | 6.7 ± 1.3 mM a,d | 0.61 ± 0.05 e |
Apigenin | 3.4 ± 0.5 mM a,d | 1.49 ± 0.09 e |
Catechin | 13.6 ± 1.8 mM a,d | 0.42 ± 0.03 e |
l-Ascorbic acid | 14.0 ± 1.9 mM a,d | 0.83 ± 0.09 e |
Glutathione | 19.7 ± 3.2 mM b,d | n.d. |
α-tocopherol | 1.1 ± 0.1 mM a,d | 3.43 ± 0.25 e |
β-carotene | 7.8 ± 0.9 μM b,d | n.d. |
LOX–FL Reaction | ||
Trolox | 5 ± 0.6 μM a; 26 ± 2 μM b,d | 1.00 |
Albumin | 25 ± 2 μM b,d | 1.04 ± 0.08 e,f |
Bilirubin | 7.6 ± 0.6 μM b,d | 3.43 ± 0.28 e,f |
l-Ascorbic acid | 1360 ± 10 μM b,d | 0.02 ± 0.002 e,f |
Uric acid | 40 ± 2 μM b,d | 0.66 ± 0.04 e,f |
a Ki values were obtained by measuring reaction rates at various RNO, FL, and antioxidant concentrations; b IC50 values represent the antioxidant concentration capable of halving the rates of LOX/RNO (or LOX-mediated oxodiene generation in the case of glutathione) or LOX–FL reactions; c ratio between the gradient of the plot reporting the decrease of the rate (%) of LOX/RNO or LOX–FL reactions; d mean value minus standard deviation (n = 4); e mean value minus standard error (n = 4); f unpublished data; n.d. Pastore et al. 25 and Soccio et al. 26 were used as inspiration.
Trolox, a noncompetitive and competitive inhibitor of LOX/RNO and LOX–FL reactions with Ki values of about 7 mM and 5 M, respectively, was discovered among different antioxidant compounds 25,26. In addition, for the LOX–FL reaction, an IC50 of 26 M was discovered. With the exception of -carotene, the LOX/RNO reaction was found to be less sensitive to single pure antioxidants than LOX–FL, as indicated by millimolar and micromolar Ki or IC50 values obtained, respectively. All other antioxidants studied showed a linear dependency of inhibition ( percent ) of the reaction rate on compound concentration, similar to Trolox. In light of this, the efficacy of each compound in relation to Trolox was calculated (Table 2, column Antioxidant/Trolox) by dividing the gradient of the plot relative to the antioxidant by the gradient of the plot relative to Trolox. Resveratrol, apigenin, and -tocopherol had higher LOX/RNO activity than Trolox, ferulic and L-ascorbic acids had comparable activity, and gallic acid and catechin had lower activity. In the LOX–FL reaction, bilirubin was more active than Trolox, uric and L-ascorbic acids had lower activity, and albumin had equal activity.
Suitability of the LOX-1-Based Assays to Assess AC of Plant Food Extracts and Blood Samples
Assessment of AC of Plant Food Extracts
LOX/RNO and LOX–FL approaches have been used to assess the AC of a wide range of plant food matrices, in addition to pure antioxidant compounds. Their results were often compared to those of two well-known AC assays, such as ORAC and TEAC, which measure mainly the scavenging ability against peroxyl and ABTS+ radicals, respectively, using SET and HAT mechanisms (see also Table 1). Table 3 summarises the key findings.
Table: Antioxidant Capacities of different plant samples as evaluated by LOX/RNO and/or LOX–FL methods in comparison with TEAC and ORAC assays.
Plant Matrices and Derived Products | Extract | AC (µmol Trolox eq./g Dry Weight) | Ref. | ||
LOX/RNO and/or LOX–FL a | ORAC | TEAC | |||
Durum wheat whole grains (Triticum durum Desf., cv. Simeto) | H | 102 ± 4.8 b |
| 3.00 ± 0.05 b | 28 |
L | 46.3 ± 1.1 b |
| 0.34 ± 0.01 b | ||
FSP | 41.1 ± 9.7 b |
|
| ||
IBP | 1137 ± 65 b |
| 6.23 ± 0.12 b | ||
Durum wheat whole grains (Triticum durum Desf., cv. Ofanto) | H | 116 ± 9 b | 19.3 ± 2.1 | 2.98 ± 0.09 b | 25,28 |
L | 57.3 ± 1.1 b | 2.26 ± 0.17 | 0.35 ± 0.01 b | ||
FSP | 133 ± 44 b |
|
| ||
IBP | 1336 ± 44 b | 14.7 ± 2.0 | 6.00 ± 0.20 b | ||
Durum wheat whole grains (Triticum durum Desf., cv. Adamello) | H | 106 ± 8 | 17.3 ± 1.5 | 5.13 ± 0.40 | 29 |
L | 73.3 ± 11.9 | 1.46 ± 0.06 | 0.24 ± 0.04 | ||
FSP | 47.2 ± 0.8 | 3.42 ± 0.16 | 0.89 ± 0.02 | ||
IBP | 800 ± 12 | 13.5 ± 1.5 | 6.67 ± 0.14 | ||
Wheat flour made from whole grains (Triticum aestivum L., cv. Bolero) | H | 128 ± 10 | 32.5 ± 3.5 | 5.34 ± 0.10 | 57,58 |
L | 146 ± 14 | 15.5 ± 0.9 | 0.19 ± 0.02 | ||
FSP | 54.0 ± 2.0 | 2.51 ± 0.11 | 0.97 ± 0.03 | ||
IBP | 256 ± 8 | 8.28 ± 1.07 | 3.92 ± 0.21 | ||
Einkorn beans, whole and unprocessed (Triticum monococcum L. ssp. sinskajae) | H | 115 ± 12 | 4.45 ± 0.16 | 5.61 ± 0.09 | 57,58 |
L | 210 ± 8 | 2.83 ± 0.06 | 0.58 ± 0.02 | ||
FSP | 74.2 ± 2.3 | 2.42 ± 0.12 | 0.73 ± 0.03 | ||
IBP | 1081 ± 22 | 1.93 ± 0.07 | 0.63 ± 0.01 | ||
Einkorn grains, whole and hulled (Triticum monococcum L. ssp. monococcum) | H | 72.3 ± 3.8 | 3.59 ± 0.02 | 5.76 ± 0.30 | 57,58 |
L | 148 ± 11 | 2.95 ± 0.25 | 0.40 ± 0.05 | ||
FSP | 263 ± 6 | 1.70 ± 0.01 | 0.51 ± 0.01 | ||
IBP | 2008 ± 38 | 12.0 ± 1.0 | 4.95 ± 0.17 | ||
Emmer (Triticum dicoccum Schübler, cv. Molise Colli) whole grains | H | 360 ± 65 | 54.6 ± 13.0 | 5.84 ± 0.12 | 29 |
L | 141 ± 11 | 0.88 ± 0.03 | 0.20 ± 0.01 | ||
FSP | 14.2 ± 0.7 | 4.25 ± 0.41 | 0.75 ± 0.04 | ||
IBP | 1239 ± 21 | 26.0 ± 0.4 | 1.61 ± 0.09 | ||
Spelt grains are whole grains (Triticum spelta L., cv. Altgold Rotkorn) | H | 256 ± 11 | 22.5 ± 1.7 | 5.23 ± 0.15 | 57,58 |
L | 104 ± 12 | 2.27 ± 0.02 | 0.80 ± 0.02 | ||
FSP | 186 ± 4 | 2.51 ± 0.07 | 0.66 ± 0.02 | ||
IBP | 1764 ± 25 | 5.84 ± 0.25 | 1.50 ± 0.05 | ||
Finger millet grains in their whole form (Eleusine coracana L. Gaertn.) | H | 565 ± 5 |
| 25 ± 0.1 | 59,60,61 |
Teff (Eragrostis tef (Zucc.) Trotter) whole grains | H | 256 ± 8 |
| 2 ± 0.1 | 59,60,61 |
Buckwheat grains are whole grains (Fagopyrum esculentum Moench) | H | 82 ± 9.8 |
| 16 ± 1.2 | 60,61 |
Amaranth grains, whole (Amaranthus spp.) | H | 64 ± 1 |
| 3 ± 0.1 | 59,60,61 |
Quinoa grains that are saponin-free (Chenopodium quinoa Willd., cv. Real) | H | 138 ± 11 | 37 ± 1 | 12.8 ± 0.5 | 29 |
L | 130 ± 6 | 0.38 ± 0.03 | 0.33 ± 0.03 | ||
FSP | 81 ± 4 | 5.75 ± 0.23 | 1.67 ± 0.06 | ||
IBP | 428 ± 4 | 4.89 ± 0.15 | 3.72 ± 0.17 | ||
Wheat bran from durum wheat | BW | 749 ± 50 (12.5 ± 0.7 a) f | 48 ± 4 f | 18.9 ± 1.3 f |
|
G Lisosan (nutritional supplement) | H | 1576 ± 427 (81.7 ± 4.7 a) | 123 ± 6 | 48 ± 3 | 26,30 |
L | 258 ± 2 (0.56 ± 0.06 a) | 1.3 ± 0.02 | 3.7 ± 0.78 | ||
FSP | 83 ± 2 (4.3 ± 0.4 a) | 25.6 ± 0.7 | 7.1 ± 1.2 | ||
IBP | 1294 ± 24 (7.1 ± 0.1 a) | 56.6 ± 4.8 | 29.5 ± 2.1 | ||
Fiber from an apple (Malus domestica Borkh.) | H | 333 ± 23 |
| 29 ± 2 | 60,61 |
Silverskin coffee | H | 1773 ± 108 |
| 27 ± 0.3 | 60,61 |
Pasta made from durum wheat semolina | H | 2.55 ± 0.09 a | 5.15 ± 0.55 | 2.29 ± 0.20 | 34 |
L | 0.41 ± 0.04 a | 1.11 ± 0.11 | 0.14 ± 0.02 | ||
FSP | 0.34 ± 0.02 a | 1.46 ± 0.16 | 0.20 ± 0.03 | ||
pasta made with durum wheat semolina and durum wheat bran oleoresin extract | H | 2.93 ± 0.11 a | 6.03 ± 0.49 | 2.13 ± 0.1 | 34 |
L | 1.58 ± 0.1 a | 0.86 ± 0.11 | 0.19 ± 0.003 | ||
FSP | 0.29 ± 0.01 a | 1.69 ± 0.19 | 0.19 ± 0.01 | ||
pasta made with durum wheat semolina and durum wheat bran water extract | H | 2.39 ± 0.1 a | 4.56 ± 0.25 | 2.34 ± 0.12 | 34 |
L | 0.35 ± 0.02 a | 1.13 ± 0.08 | 0.15 ± 0.01 | ||
FSP | 0.92 ± 0.03 a | 1.50 ± 0.03 | 0.29 ± 0.01 | ||
Japanese knotweed (Polygonum cuspidatum Siebold et Zucc.) root contains 98 percent food-grade resveratrol. | - | 28.2 ± 0.6 c |
| 6.44 ± 0.71 c | 31 |
Food-grade quercetin (98%) extracted from the flower buds of the Japanese pagoda tree (Sophora japonica L.). | - | 12.7 ± 0.5 c |
| 5.87 ± 0.50 c | 31 |
Catechins (50%) in food-grade form from green tea (Camellia sinensis (L.) Kuntze) leaf | - | 29.8 ± 0.7 c |
| 3.15 ± 0.45 c | 31 |
Lycopene (15%) in food-grade form from tomato (Solanum lycopersicum L.) fruit | - | 215 ± 1.5 c |
| 2.45 ± 0.30 c | 31 |
OLIPLUS® is a polyphenol-rich olive (Olea europaea L.) extract (45 percent ) | - | 24.0 ± 1.0 c |
| 1.69 ± 0.30 c | 31 |
olive oil (extra virgin) (Olea europaea L., cv. Cima di Mola) | - | 4030 ± 400 d | 17.3 ± 1.8 d | 4.4 ± 0.2 d | u.d. |
olive oil (extra virgin) cv. Coratina (Olea europaea L.) | - | 2800 ± 160 d | 26.0 ± 2.7 d | 3.9 ± 0.3 d | u.d. |
olive oil (extra virgin) (Olea europaea L., cv. Peranzana) | - | 2770 ± 10 d | 14.2 ± 0.1 d | 2.01 ± 0.07 d | u.d. |
a glass of red wine (Vitis vinifera L., cv. Negramaro) | - | 8.6 ± 0.2 a,d | 59.8 ± 5.8 d | 24.4 ± 1.3 d | u.d. |
a glass of red wine (Vitis vinifera L., cv. Nero di Troia) | - | 10.7 ± 0.3 a,d | 47.1 ± 2.8 d | 36.6 ± 0.9 d | u.d. |
a glass of red wine (Vitis vinifera L., cv. Primitivo) | - | 8.0 ± 0.3 a,d | 47.6 ± 6.0 d | 32.6 ± 1.3 d | u.d. |
Cherry (Prunus avium L., cv. Ferrovia) fruit puree | FSP | 5.1 ± 0.2 a,e | 19.1 ± 0.1 e | 6.5 ± 0.1 e | 35 |
Fruit of the peach (Prunus persica L., cv. Redhaven) | C | 155 ± 10 e | 0.33 ± 0.04 e | 0.082 ± 0.002 e | 32 |
Fruit of the peach (Prunus persica L., cv. Armking) | C | 125 ± 10 e | 0.44 ± 0.06 e | 0.068 ± 0.001 e | 32 |
Fruit of the peach (Prunus persica L., cv. Silverking) | C | 9.6 ± 3.5 e | 0.21 ± 0.04 e | 0.025 ± 0.001 e | 32 |
Fruit of the peach (Prunus persica L., cv. Caldesi 2000) | C | 10.5 ± 1.2 e | 0.11 ± 0.01 e | 0.018 ± 0.0001 e | 32 |
Fruit of the peach (Prunus persica L., cv. IFF331) | C | 9.8 ± 1.5 e | 0.13 ± 0.01 e | 0.020 ± 0.0001 e | 32 |
Fruit of the tomato (Solanum lycopersicum L.) | H | 3.79 ± 0.84 a | 64.4 ± 10.7 | 38.3 ± 4.0 | u.d. |
L | 11.25 ± 5.24 a | 41.6 ± 15.4 | 3.88 ± 0.87 | ||
Pomegranate (Punica granatum L.) fruit juice | - | 36.2 ± 5.4 a,d | 14.6 ± 3.1 d | 45.8 ± 1.5 d | u.d. |
H stands for hydrophilic extract; L stands for lipophilic extract; FSP stands for free-soluble phenolic extract; IBP stands for insoluble-bound phenolic extract; C stands for carotenoid-enriched extract; u.d. stands for unpublished results. Without extracting the samples, olive oil, red wine, and pomegranate juice were analysed. a mol Trolox eq./mg fresh weight; b mean value (SE) of four whole grain samples collected from plants subjected to various growing conditions (for information, see Laus et al. 28); c mol Trolox eq./mL; d mol Trolox eq./g fresh weight
To begin, the LOX/RNO assay was used to dissect AC of whole durum wheat grains 25,28. Cereal whole grains, in particular, have attracted increasing attention in recent decades as a result of their unusual and peculiar content in biologically active compounds, which are thought to be responsible for some of the health benefits associated with daily grain intake, such as defence against chronic diseases and diet-related disorders (for a recent review, see Masisi et al. 62). The LOX/RNO process, in particular, revealed significant differences between the antioxidant-enriched extracts from durum wheat grains, with the highest AC values in the phenolic fraction linked to insoluble cell wall polymers, which is known as the key antioxidant component in cereal grains 63. Furthermore, when it came to the TEAC assay, the LOX/RNO system was able to distinguish between samples collected from different cultivars and under different experimental conditions (including different fertilisation and irrigation treatments), as well as provide accurate AC information that was highly correlated to antioxidant compound content 28. LOX/RNO assay has also been used to test AC of various extracts obtained from grains of emmer 29, bread wheat, spelt, hulled and naked einkorn 57,58 in addition to durum wheat. Interestingly, unlike TEAC and ORAC assays, LOX/RNO was able to reveal significant differences in the balance of free and bound grain antioxidants among the cereal species studied, with the hulled cereal species clearly superior for both AC of insoluble-bound phenolic portion and AC of freely solvent-soluble antioxidants. This may indicate that antioxidants have very different potential effects on consumer health, with free antioxidants having a stronger systemic safe activity and bound antioxidants having a useful activity at the terminal intestine stage 64,65,66. Antioxidant properties of teff and finger millet grains were also measured by the LOX/RNO method 59,60,61, as well as AC of the pseudocereals quinoa 29, amaranth and buckwheat 59,60,61. In the case of quinoa, LOX/RNO was able to unearth very interesting antioxidant properties, possibly related to health benefits, in comparison to the other compared AC assays. LOX/RNO found that the freely soluble and more easily accessible antioxidant fraction of quinoa seeds has a much higher AC than whole grains of the commonly consumed cereals durum wheat and emmer, piqueing interest in quinoa-derived products not only for celiac diet but also for general population nutrition 29.
Both LOX-1-based assays were used to investigate the AC of certain cereal-grain-derived products. Both the LOX/RNO 30 and LOX–FL methods 26 were used to investigate the antioxidant properties of Lisosan G, an antioxidant-rich dietary supplement made from lysed fine bran and germ of organic wheat grains with well-documented bioactivity 67,68,69,70. Both LOX-1-based methods reported a remarkable total AC of Lisosan G, which is consistent with the strong antioxidant properties of fermented whole grain products due to the improved content of water-soluble compounds 62. It was 3- to 10-fold higher than that measured for various cereal species, and it is primarily due to a highly active freely water-extractable antioxidant portion, as well as insoluble-bound phenols 26,30.
In a hydrophilic/phenolic antioxidant-rich extract obtained by ultrasound-assisted water extraction of durum wheat bran, very high AC values were calculated by both LOX/RNO and LOX–FL assays in a cereal-grain-derived product (bran water extract, BW extract). In addition, LOX–FL assay was used to dissect AC of two cooked semolina pastas, supplemented with Bran Oleoresin (BO) extract rich in lipophilic (tocochromanols, carotenoids) antioxidants and with BW extract, in comparison with a non-supplemented pasta 34. Interestingly, LOX–FL found the highest AC values in the free-soluble phenolic and lipophilic extracts of BW and BO pastas, respectively, in accordance with the separate supplementation of BW and BO pastas, while ORAC and TEAC 34 found the opposite performance.
It should also be noted that LOX/RNO proved to be an excellent tool for determining AC in food-grade preparations derived from a variety of plant sources and highly enriched in either lycopene or different phenolic compounds, including the flavonoids catechins and quercetin, as well as the non-flavonoid resveratrol and tyrosol/hydroxytyrosol/oleuropeine mixture (OLIPLUS®) 31
LOX-1-based assays also performed well in determining the AC of fruits and vegetables. In particular, in carotenoid-enriched extracts from peach fruits, the LOX/RNO method measured AC values highly related to carotenoid content, and pointed out a much higher differences among yellow- and white-fleshed varieties than ORAC and TEAC methods 32. The LOX–FL assay was also used to test the free-soluble phenolic portion obtained from cherry puree 35, as well as pomegranate fruit juice and hydrophilic and lipophilic fractions from tomato fruit. Finally, the LOX–FL assay has been used to evaluate the antioxidant properties of extremely complex matrices like extra-virgin olive oil and red wine.
In conclusion, LOX-1-based assays have proven to be useful tools for determining the antioxidant capacity of a wide range of plant foods, demonstrating the ability to distinguish between different samples and provide accurate AC information correlated to antioxidant content and likely related to health benefits.
Assessment of AC of Blood Samples
In addition to being used for AC measurements of food extracts, as previously mentioned (Section 2), the LOX–FL assay has the added dimension of being an appropriate instrument for analysis of biological samples, such as human blood samples 26. In reality, the LOX–FL method can be used on both human blood serum and plasma, while the LOX/RNO assay requires an excessive amount of both biological fluids. When plasma is obtained using EDTA or citrate as anticoagulants, however, caution must be exercised because these anticoagulants can chelate iron, which is needed for LOX operation. In light of this, the LOX–FL assay has been favoured for ex vivo AC evaluation of human blood serum. The LOX–FL reaction has been used to test a significant number of serum samples to date. Within a very broad range of 0.8 to 2 mol Trolox eq./mL of serum, a normal distribution of AC data was collected (Figure 3). Interestingly, the LOX–FL assay may play an important role in the second stage of investigation, which involves AC analysis of serum/plasma after food intake, for the property to be extended to both in vitro and ex vivo AC measurements. The high output of the LOX–FL system in both in vitro and ex vivo tests has the benefit of allowing for an integrated AC assessment of food antioxidants: through using the same assay, it is possible to determine if foods with high AC can provide a real benefit after ingestion.
 Figure 3 Distribution of serum Antioxidant Capacity values obtained by LOX–FL assay. Serum AC values were obtained from 92 subjects after an overnight fast.
Figure 3 Distribution of serum Antioxidant Capacity values obtained by LOX–FL assay. Serum AC values were obtained from 92 subjects after an overnight fast.
Evaluation of Synergistic Effects Among Antioxidants Using LOX-1-Based Assays
The ability of LOX-1-based AC assays to reveal synergism between antioxidant compounds has been demonstrated. In contrast to ORAC and TEAC assays, Table 4 shows the results of various experiments aimed at evaluating the capacity of both LOX/RNO and LOX–FL reactions to illustrate antioxidant synergistic interactions.
In comparison to ORAC and TEAC assays, Table 4 shows synergism among antioxidant compounds evaluated by LOX/RNO and LOX–FL methods.
| LOX/RNO a or LOX–FL b | ORAC | TEAC | Ref. |
Among Different Pure Compounds (% Change) c | ||||
Resveratrol, quercetin, OLIPLUS®, catechin, and lycopene are all food-grade extracts. | +570 ***,a | - | +30 ** | 31 |
Ascorbic acid, Trolox, bilirubin, uric acid, and albumin are all mixed together. | +74 ***,b | +12 n.s. | - | 26 |
| Among different extracts (% change) c | |||
phenolic extracts from durum wheat whole flour that are hydrophilic, lipophilic, and insoluble-bound | +108 **,a | +39 * | −32 * | 25 |
| Among phenols in the same extract (time fold change) d | |||
Phenols from whole durum wheat flour that are insoluble | 410 a | 2.8 | 0.89 | 28 |
| Among human serum and free-soluble phenols (% change) c | |||
Human blood serum and Lisosan G's free-soluble phenolic extract | +124 ***,b | +16 * | - | 26 |
a a LOX/RNO evaluation; b a LOX–FL evaluation; c a change ( percent ) in the AC of the mix compared to the amount of the AC of each individual compound (or extract); d a time fold change in the IC50 values of pure ferulic acid compared to that of phenolic extract. n.s. = not significant, according to the Student's t-test; *, p 0.05; **, p 0.01; ***, p 0.001; n.s. = not significant, according to the Student's t-test.
By comparing the AC of a mixture of compounds to the amount of the AC of individual compounds, the LOX/RNO method outperformed TEAC in assessing synergistic interactions among food-grade preparations rich in phenolic compounds such as catechins, quercetin, resveratrol, and tyrosol/hydroxytyrosol, as well as carotenoids like lycopene 31. Similarly, the LOX–FL assay was able to detect synergistic effects among the most common antioxidants found in human blood serum, such as ascorbic and uric acids, albumin, bilirubin, and the vitamin E analogue Trolox. In particular, when compared to ORAC, the ability to illustrate synergistic interactions was found to be around 6 times higher 26.
Furthermore, the LOX/RNO assay detected a three-fold higher synergism among hydrophilic, phenolic, and lipophilic antioxidant compounds extracted from durum wheat whole flour than the ORAC assay, whereas the TEAC method was ineffective in the same experimental conditions 25.
By measuring interactions within antioxidant compounds in the same type of extract, the performance of the LOX/RNO reaction in detecting synergism was also evaluated. The inhibition of the reaction rate by phenolic compounds extracted from durum wheat whole flour was compared to the inhibition exerted by a high-purity ferulic acid preparation. Using this protocol, LOX/RNO displayed a 400-fold higher inhibition effectiveness of phenolic mixture than pure compound, suggesting a very strong synergistic activity of phenols within the extract, while the same comparison for ORAC and TEAC assays yielded inhibition effectiveness of only about 2.8 and 0.89 28.
Finally, the LOX–FL method was used to calculate an eight-fold higher synergism between human serum and food phenolic antioxidants obtained from the dietary supplement Lisosan G than the ORAC assay 26.
In conclusion, LOX-1-based assays are far more effective than other well-known AC assays in revealing synergism among antioxidant compounds, most likely due to their ability to simultaneously detect several antioxidant functions. This has been demonstrated using a variety of experimental procedures involving a mixture of various concentrations of antioxidants with varying chemical–physical properties and derived from a variety of plant sources. Since synergistic interactions among food antioxidant compounds are thought to play a critical role in the health-promoting effects of some plant foods 63, the ability to detect synergism is a very important property of an AC assay.
Overall, the benefits of both LOX-1-based assays can be summarised as follows: To begin, the LOX/RNO and LOX–FL methods use protocols that are I technically easy and require readily available instrumentation, (ii) capable of ensuring good reproducibility of findings, and (iii) easily applicable to single pure antioxidant compounds, food extracts, both fat-soluble and water-soluble, as well as biological samples (plasma, and serum). Furthermore, these assays allow measurements to be carried out in experimental conditions that are similar to those that occur in vivo. In fact, in LOX-1-based reactions, I more than one oxidant species with biological significance is involved, (ii) oxidant–antioxidant competition is assessed at low oxygen concentrations, and (iii) it occurs at the surface of a biological macromolecule (the LOX enzyme) rather than in the bulk phase of the reaction mixture. Furthermore, LOX-1-based assays can test multiple antioxidant mechanisms at the same time, as well as synergistic interactions between antioxidant compounds. In the light of all these properties, LOX-1-based assays may provide a reliable, advanced and comprehensive AC information of natural plant products, potentially more reflecting their in vivo biological effects compared to the most widespread AC assays. Interestingly, the main requirements/criteria proposed by Prior et al. 1 for standardization of AC determination may be considered sufficiently satisfied by both LOX–RNO and LOX–FL methods.
4.1.5 Evaluation of Human Blood Antioxidant Status after Food Intake
Antioxidant/Oxidant Balance (AOB) Approach and its Application in Short-Term Studies
It should be noted that in vitro AC testing may only allow for merceological characterization of plant food commodities and cannot be linked to biological effects of dietary bioactive compounds after ingestion 7,21. In reality, in vitro AC analysis of food extracts does not provide information on dietary phytochemical bioavailability, that is, the fraction of ingested antioxidant compound that reaches the systemic circulation, as well as in vivo stability, tissue retention, and in situ reactivity. The United States Department of Agriculture (USDA) removed its ORAC database for selected foods from its nutrient data laboratory website 22 in 2012 as a result of these factors.
A new method has been suggested to gain more physiologically meaningful knowledge about potential health effects of foods by combining in vitro AC examination of foods with an ex vivo study of blood antioxidant status after food consumption. The bioavailability of dietary phytochemicals as well as the metabolism of food antioxidants, i.e., the potential transformations of phytochemicals through gut microbiota and intestinal and hepatic metabolism, may be taken into account when assessing serum/plasma AC after food intake 71. This may provide a more comprehensive picture of the true impact of food antioxidant assumptions on blood antioxidant status, which goes beyond the AC of the consumed food and cannot be determined solely by in vitro AC measurements.
Unfortunately, this strategy may have some flaws, leading to some contentious outcomes. In fact, blood AC measurements after ingestion of antioxidant-rich foods have shown contrasting results, with only a small increase 72,73, no effect 74,75, and in some cases, even a decrease 35,75. It's also worth noting that some studies found no improvement in AC followed by a decrease in serum/plasma oxidation levels 35,76. These results indicate that blood AC tests alone are unsuitable for determining food antioxidant efficacy, and that changes in serum oxidative status must also be considered. In light of this, an improvement in the calculation of serum antioxidant status was attempted by proposing a novel parameter called "Antioxidant/Oxidant Balance" as a means of overcoming this flaw (AOB). It is obtained as a ratio between serum AC and serum oxidant status, evaluated as "Peroxide Level" AOB = AC/Peroxide Level (PxL) 34. It includes evaluation of both antioxidant and oxidant status of blood after food intake and is obtained as a ratio between serum AC and serum oxidant status, evaluated as "Peroxide Level" AOB = AC/Peroxide Level (PxL) 34. It should be noted that the results of PxL's serum hydroperoxides assay 34 are strongly associated with those of the oxidised low-density lipoproteins assay 34.
For the first time, the efficacy of the novel AOB method in illustrating improvements in serum antioxidant status after food intake was evaluated by comparing the short-term (up to 4 h) effects of two antioxidant-enriched pastas, supplemented with bran oleoresin (BO) and bran water (BW) extracts, respectively, to a non-supplemented reference (R) pasta 34. Initially, the efficiency of the AOB parameter was verified following the consumption of two foods that are supposed to have the opposite effect on serum AC. They included Lisosan G, a dietary supplement with a high in vitro AC of 26,30 (see also Table 3) and glucose, which is considered to have a pro-oxidant effect 77,78. AOB profiles obtained within 4 hours after Lisosan G and glucose assumption by measuring ACLOX–FL/PxL, ACORAC/PxL, or ACTEAC/PxL ratios are compared to AC and PxL profiles in Laus et al. 34. Although serum AC measurements varied between assays 79, AOB profiles measured as ACLOX–FL/PxL, ACORAC/PxL, or ACTEAC/PxL all agreed on a significant improvement in serum antioxidant status after Lisosan G intake, as well as a significant decrease due to glucose ingestion 34. In the case of pastas, all AOB profiles showed a decrease in serum antioxidant status after R pasta consumption, comparable to glucose; a decreasing trend was also seen after R pasta consumption combined with Lisosan G 34. In a similar way to Lisosan G 34, BO pasta intake resulted in a substantial increase in AOBLOX–FL, as well as AOBORAC or AOBTEAC. BW pasta, on the other hand, was not only unable to have a beneficial effect on serum, but it also had an oxidative effect, as did R pasta and glucose.
A quantification involving the measurement of region under AOB profiles vs time was proposed to promote the comparison of short-term effects of food intake on serum AOB. AOB-Area and AOB-Index are two AOB-derived parameters that have been suggested 34. As shown in Figure 4, AOB-Area is calculated as a percentage change in the area under the serum AOB profile vs. the period of the tested food in relation to the basal area, i.e., the area below the fasting value. The AOB-Index is measured as a percentage change in the area under the AOB profile compared to the area obtained after 50 grammes of glucose intake, thus accounting for the starch portion of pasta (in terms of glucose released from starch digestion).
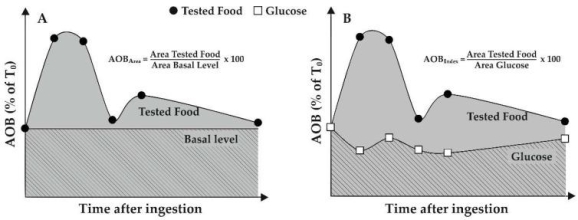
After food intake, the AOB-Area and AOB-Index are calculated. The authors present hypothetical AOB profiles in relation to a generic diet and glucose consumption. (A) AOB-Area is measured as a percentage of the area under the serum AOB (expressed as a percentage of T0 value, i.e., value obtained before food intake) profile vs time (grey coloured area) and the area below the baseline (crossed area), i.e., area below the baseline value (T0). (B) The AOB-Index is measured as a percentage difference between the area under the AOB profile and the area obtained after consuming 50 grammes of glucose (crossed area).
Figure 5 compares serum AOB-Area (A–C) and AOB-Index (A'–C') after consumption of BO, BW, and R pastas to that after consumption of Lisosan G, glucose, and R pasta added with Lisosan G. AOB-Area clearly demonstrated the superiority of the antioxidant-rich supplement Lisosan G over the pro-oxidant glucose, regardless of the assay used to calculate AC. AOB-AreaLOX–FL or AOB-AreaORAC and AOB-AreaTEAC, respectively, attributed to BO pasta a significant antioxidant effect on serum, comparable to or even higher than that of Lisosan G. BW pasta, on the other hand, had a pro-oxidant effect on serum, as shown by AOB-Area values of ACLOX–FL/PxL, ACORAC/PxL, or ACTEAC/PxL, which were equal to or even lower than glucose. When R pasta was combined with Lisosan G, a behaviour similar to glucose was observed. AOB-Area determined as ACLOX–FL/PxL and ACORAC/PxL or ACTEAC/PxL, respectively, found a worsening equal to and slightly higher than glucose for the non-supplemented R pasta, as predicted. The different actions of BW and BO pastas indicated that hydrophilic/phenolic antioxidants were ineffective in compensating serum oxidation due to glucose release from starch digestion under the experimental conditions used, and that lipophilic compounds were capable of both counteracting the negative effects of starch/glucose and improving serum antioxidant status 34.
After ingestion of various foods in seven subjects, the AOB-Area ((A), (B), (C)) and AOB-Index ((A'), (B'), (C') of serum were evaluated as ACLOX–FL/PxL (AOBLOX–FL), ACORAC/PxL (AOBORAC), and ACTEAC/PxL (AOBTEAC) ratios. Serving sizes for pastas were 70 g, Lisosan G was 20 g (fresh weight), and glucose was 50 g. Each tested food's AOB-Area and AOB-Index values reflect areas under the AOB profile vs. time (from 0 to 240 minutes), expressed as ( percent ) of basal area and ( percent ) of area relative to glucose intake, respectively (see Figure 4). The data is presented as a mean (n = 7 subjects). According to the Duncan's test, different letters within the same graph suggest major variations at a p-value of 0.05. BO pasta: bran oleoresin-supplemented pasta; R pasta: reference pasta; BW pasta: bran water extract-supplemented pasta The data from Laus et al. 34 has been adequately re-elaborated.
The AOB-Index data basically validated the AOB-Area data, but it was able to amplify the changes (Figure 5A'–C'). In fact, in addition to confirming the similar actions of BW and R pastas, AOB-IndexLOX–FL revealed that BO pasta and Lisosan G had a remarkable beneficial effect on serum antioxidant status of up to +70% and +65%, respectively, when compared to R pasta. AOB-IndexTEAC and AOB-IndexLOX–FL both noted a major increase in R pasta consumed with Lisosan G as compared to R pasta alone.
When AOB responses are compared to those obtained via ex vivo serum AC analysis, some interesting findings emerge. In fact, only the LOX–FL assay was able to distinguish between Lisosan G and glucose when measuring serum AC, while ORAC and TEAC failed 34,79. Furthermore, only the LOX–FL system detected a small increase in serum AC after BO pasta consumption compared to both BW and R pastas, while TEAC and ORAC found no difference 34,79. If in vitro AC measurements are taken into account, the LOX–FL, ORAC, and TEAC methods agree only in highlighting Lisosan G's remarkable AC, but produce different results on cooked pastas 34. (see also Table 3). Overall, these results support the suitability and reliability of the AOB method, which can detect short-term changes in blood antioxidant status after eating antioxidant-rich foods that cannot be predicted by ex vivo AC analysis or in vitro food measurements. Surprisingly, using the AOB parameter allows ORAC and TEAC assays to work again. In light of these findings, the AOB parameter tends to be a more effective method for evaluating the effects of food intake on serum antioxidant status than the AC assessment.
Finally, the strong antioxidant properties of the dietary supplement Lisosan G, as well as the pro-oxidant impact of glucose intake, were verified using this novel approach to short-term food consumption evaluation. Interestingly, the real health effects of two antioxidant-supplemented pastas were verified using the same process. In particular, AOB has allowed highlighting the pasta supplemented with durum wheat bran oleoresin extract's strong efficacy in enhancing serum antioxidant status, as well as excluding a potential biological effect of the other putative functional pasta, added with bran phenolic compounds 34.
AOB Approach in Long-Term Studies
Two long-term small pilot studies 35 were used to test the reliability of the AOB approach. For the first sample, eight participants ate 250 g of cherry puree per day for three weeks. In the second study, 11 participants were enlisted to take part in the “Med-Food Anticancer Program (MFAP)” nutrition education intervention, which included promoting a healthy Mediterranean diet that included consumption of many antioxidant-rich plant foods to achieve a typical daily antioxidant intake of about 50% higher 35. AC tests were unable to detect a rise in serum AC; in both experiments, a 7–9% significant decrease was estimated using LOX–FL and ORAC methods. PxL, on the other hand, showed a much more pronounced (about 20%) significant decrease 35. Surprisingly, when the study was switched from AC to AOB, both long-term studies were able to show a significant increase in serum antioxidant status (ranging from around 20% to 40%) 35. This demonstrates the new method's reliability in highlighting the anticipated positive improvements in serum antioxidant status as a result of long-term dietary consumption of antioxidant-rich plant foods. Surprisingly, the AOB approach was able to discover the health benefits of any form of dietary antioxidant administered.
Overall, the AOB method tends to be a more effective strategy than relying solely on in vitro and ex vivo AC determinations. Surprisingly, the AOB parameter yields similar results regardless of which assay is used to calculate AC, though the novel LOX–FL method performed better, owing to its ability to provide more biologically relevant AC details. This high performance of the AOB method is due to the simultaneous assessment of AC and PxL, which allows for the accounting of both AC and the AC fraction not explicitly measureable as consumed to compensate for serum oxidation 34,35, and can help to resolve some misleading results obtained by ex vivo serum AC determination alone. It's still unclear if a positive impact on blood antioxidant status can be linked to a health benefit. However, since maintaining blood physiological antioxidant status is needed to prevent aging-related endothelial dysfunction and subsequent cardiovascular disease 80, AOB can provide biologically relevant details.
It should also be noted that the AOB method has recently been used as a biomarker of blood antioxidant status in ewes fed Ascophyllum nodosum and flaxseed under high ambient temperature 81 and in sheep suffering from foot rot and treated with ozone therapy 82, demonstrating its versatility and applicability in both human and animal studies.
4.1.6 Conclusions
Methodological approaches can play an important role in evaluating the potential health benefits of dietary phytochemicals. In fact, proper methodology selection is needed to avoid inaccurate and misleading data interpretation about antioxidant properties of foods. In this paper, a novel advanced analytical approach to this problem is discussed. It combines I in vitro AC measurements of food extracts using novel assays based on soybean LOX-1 reactions capable of producing many physiological radical species in experimental conditions similar to cellular ones and revealing different antioxidant attributes, and (ii) ex vivo analysis of blood serum after food consumption, involving both the AC and peroxidation level determination. To summarise, this combined in vitro/ex vivo study of antioxidant properties of foods using LOX-1-based methodologies appears to be a high-performing tool capable of providing more integrated and trustworthy knowledge about the possible health benefit of dietary antioxidants than AC analysis alone.
The image that emerges from this advanced approach can be further enhanced by assessing phytochemical biological effects at the cellular/subcellular level.
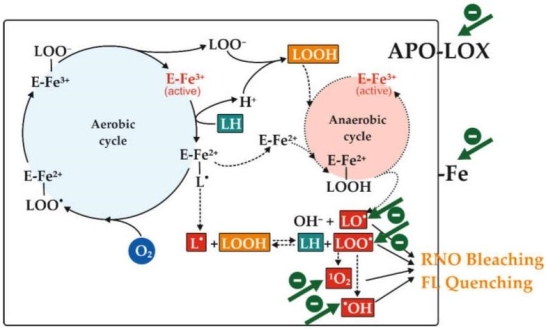
Scheme 1
The soybean lipoxygenase (LOX)-1 isoenzyme catalyses aerobic and anaerobic reactions involved in 4-nitroso-N,N-dimethylaniline (RNO) bleaching or fluorescein (FL) quenching. The aerobic cycle (continuous line) of LOX-1-catalyzed dioxygenation of linoleic acid (LH, highlighted in teal) to 13-hydroperoxy-linoleate (LOOH, highlighted in orange) is shown, as well as the secondary anaerobic cycle (dotted line) involving LOX-1 under low oxygen conditions and producing some reactive species (highlighted in red). RNO bleaching or FL quenching can occur as a result of these factors. Antioxidant compounds can inhibit this through a variety of mechanisms (green arrows), including scavenging one or more radical species, chelating or reducing iron ions, and inhibiting the apoenzyme. L∙, linoleate alkylic radical; LO∙, linoleate alkoxyl radical; LOO∙, linoleate peroxyl radical; 1O2, singlet oxygen; ∙OH, hydroxyl radical. The non-heme iron atom that plays a key role in LOX-1 catalysis is denoted by the letter "Fe." Pastore et al. 25. Adapted from Pastore et al.
4.2.1 Hoffmann’s exhaustive methylation:
Exhaustive methylation is the process of alkylation with methyl groups that continues until all possible methylations are achieved.
Prior to E2 elimination, exhaustive methylation (shown in red) transforms an amine to an alkylammonium salt in the Hofmann elimination reaction chain.
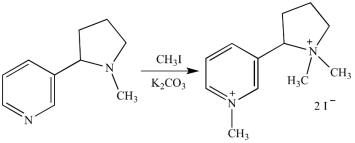
In natural product structural analysis, exhaustive methylation has been used. If one mole of an unknown alkaloid (in this case, nicotine) reacts with two moles of methyl iodide, we can deduce that the alkaloid has two methylation sites.
4.2.2 Hoffmann Elimination:
Hofmann eliminations are 4o-ammonium salt reduction reactions. Since the counter anion in most 4o-ammonium salts is halide, it is frequently replaced by the more simple hydroxide ion via silver hydroxide reaction (or silver oxide). To impact the E2-like removal of a 3o-amine, the resulting hydroxide salt must be heated (100 - 200 oC). A typical Hofmann elimination is shown in Example #1 below. As previously mentioned when looking at alkyl halide elimination reactions, one of the alkyl substituents on nitrogen must have one or more beta-hydrogens for an elimination to occur.
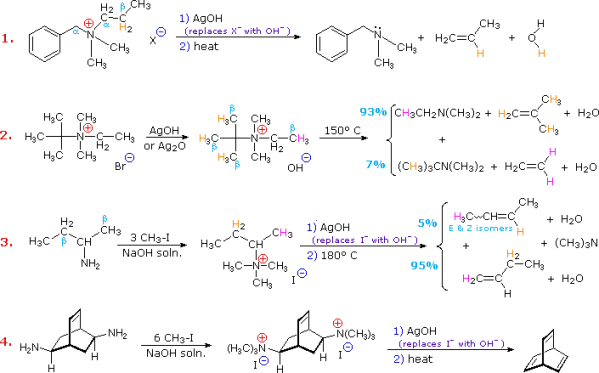
Two of the alkyl substituents on nitrogen in Example #2 have beta-hydrogens, all of which are on methyl groups (colored orange & magenta). The alkene with the more strongly substituted double bond is the main product of the removal, reflecting not only the 3:1 numerical advantage of certain beta-hydrogens, but also the greater stability of the double bond. Example #3 highlights two main aspects of the Hofmann elimination:
1. Extensive alkylation, typically with methyl iodide, converts simple amines to the necessary 4o-ammonium salts (methyl has no beta-hydrogens and cannot compete in the elimination reaction). In example #4, exhaustive methylation is demonstrated once more.
2. When an alkyl group has two sets of beta-hydrogens available for removal (coloured orange and magenta here), the alkene isomer with the less substituted double bond is always the main product.
The Hofmann Rule refers to the tendency of Hofmann eliminations to produce the less-substituted double bond isomer, which contrasts sharply with the Zaitsev Rule for dehydrohalogenations and dehydrations. The Hofmann Rule does not apply when other triggering groups, such as phenyl or carbonyl, are present. When 2-amino-1-phenylpropane is handled in the manner described in Example #3, the result is primarily 1-phenylpropene (E & Z-isomers).
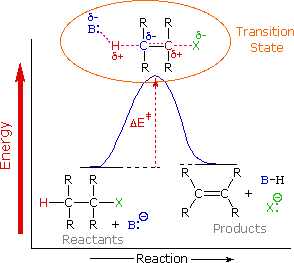
It's important to reexamine the existence of the E2 transition state, which was first identified for dehydrohalogenation, to understand why base-induced removal of 4o-ammonium salts behaves differently than that of alkyl halides. On the right, the energy diagram for a single-step bimolecular E2 process is repeated. E2 reactions have a less well-defined transition state than SN2 reactions. More bonds are breaking and forming, with the probability of a continuum of states where the degree of C–H and C–X bond breaking and C=C bond formation varies. The transition state resembles that of an E1 reaction if the bond to the leaving group (X) is significantly broken relative to the other bond changes (initial ionisation followed by a fast second step). On the other hand, if the beta-hydrogens' acidity is increased, significant C–H bond breaking can occur before the other bonds are affected. For most simple alkyl halides, it was necessary to imagine a balanced transition state in which all bonds changed at the same time. The Zaitsev Rule was compatible with such a model.
When the leaving group X has a positive charge, as it does in the 4o-ammonium compounds discussed here, the inductive effect of the charge increases the acidity of both the alpha and beta-hydrogens. Furthermore, since the 4o-ammonium substituent is far larger than a halide or hydroxyl group, it can disrupt the conformations that substituted beta-carbons may take. It appears that a combination of these factors favours base attack on the beta-hydrogens with the least substituted (least hindered and acidic) group of substituents. Many Hofmann eliminations have the preferred anti orientation of the leaving group and beta-hydrogen, as seen in dehydrohalogenation; however, syn-elimination is also normal, likely due to the attraction of opposite charges orienting the hydroxide base near the 4o-ammonium leaving group.
In the diagram below, three more examples of the Hofmann elimination are given. In two ways, Example #1 is intriguing. For starters, it produces a 4o-ammonium halide salt in a different way than exhaustive methylation. Second, prior to elimination, this salt is not converted to its hydroxide analogue. The volatile hydrocarbon substance is isolated by distillation after a concentrated aqueous solution of the halide salt is lowered into a refluxing sodium hydroxide solution.
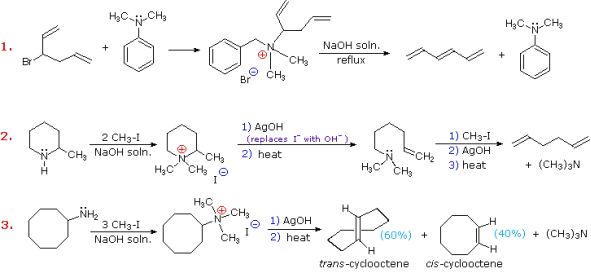
Example #2 exemplifies a key feature of the Hofmann elimination. If the nitrogen atom is part of a ring, this removal technique does not extract the nitrogen as a separate 3o-amine product in a single application. A second Hofmann elimination is needed to separate the nitrogen feature from the molecule. Indeed, if the nitrogen atom was part of two rings (fused or spiro), it would take three Hofmann eliminations to separate the nitrogen from the remaining molecular structure.
The less stable trans-cyclooctene is the main product in Example #3, which is followed by the cis-isomer. Since the cis-cycloalkene will be produced by an anti-E2-transition state, the trans-isomer must be produced by a syn-elimination. A syn-elimination may also generate the cis-cyclooctene released in this reaction. The structure of cyclooctane is conformationally complex. There are a number of puckered conformations that escape angle strain, and one of the most stable is shown on the right. Many of these conformers have several eclipsed bonds, and transannular hydrogen crowding is inevitable. Since the trimethylammonium substituent is wide (roughly the same size as tert-butyl), it will most likely take on an equatorial orientation to prevent steric crowding. An axial-like orientation of this bulky group is likely needed for an anti-E2 transition state, making this an unfavourable direction.
4.3.1 Interactions between nicotine and drugs of abuse:
Introduction
In the United States, nicotine-containing products are among the most commonly used addictive drugs. According to the most recent Substance Abuse and Mental Health Services Administration (SAMHSA) survey, 67 million people (nearly 25% of the population) use tobacco. Despite the fact that the number of people who use tobacco products has declined over the last decade, the use of modern products like nicotine-containing e-cigarettes and other electronic nicotine delivery systems (ENDS) has skyrocketed. In addition to the serious problem of nicotine addiction in the general population, cigarette smoking among people who have substance use disorders is a specific public health issue that has received insufficient attention, particularly in preclinical research. Since nicotine is a legally available drug with relatively mild psychoactive effects, the full extent of nicotine polysubstance usage is likely underreported. Even so, nicotine is frequently mentioned as a portion of the most common polysubstance combinations. In a European study of school students, nearly three-quarters (73%) of polysubstance users reported using both alcohol and cigarettes in the previous month, and one-fifth of those surveyed reported using cannabis (9-tetrahydrocannabinol; THC) concurrently with alcohol and/or nicotine.
The societal consequences of nicotine polysubstance usage are a significant source of concern. Chronic health issues linked to nicotine addiction are one of the leading causes of preventable death in the United States (e.g., 480,000 deaths per year). Tobacco use results in annual economic costs of $289 billion for cancer, cardiovascular disease, and other health risks. Furthermore, the health risks associated with medication combinations greatly outweigh the risks associated with any drug alone. As a result, polysubstance consumers often report, and laboratory tests confirm, that mixing multiple substances enhances the use of one or both of them (e.g., cocaine, d-amphetamine, heroin, and alcohol). This increases the urgency of the problem and the need for strategies to control and/or counteract the effects of this type of polysubstance abuse. This can lead to accelerated development of dependency or increased incidences of acute or long-term toxicity – reduced liver or kidney function, damage to the cardiovascular or respiratory systems, etc.– and emphasises the urgency of the problem and the need for strategies to manage and/or counteract the effects of this type of polysubstance abuse.
The interactions of nicotine with alcohol, stimulants (e.g., cocaine, amphetamines), opioids (e.g., morphine, heroin, etc.) and THC are the subject of this study. Work in some of these areas is limited, as should be obvious. The studies discussed below will illustrate the current understanding of how these various groups of abused drugs could interact with nicotine on behavioural, physiological, and pharmacological indices that may be important in maintaining co-use of either or both substances, while not intended to be exhaustive. There are some ideas for potential research areas and knowledge gaps discussed.
It is widely acknowledged that nearly 80% of nicotine-dependent adult smokers began smoking during their teenage years. Furthermore, teens who smoke cigarettes are seven times more likely than those who do not to use an illegal drug. Similarly, sex/gender differences can play a role in determining behavioural and/or neurochemical responses to drugs of abuse, as well as drug use patterns. A number of recent reviews have looked at the effects of nicotine/tobacco or other substance use during crucial developmental phases (prenatal or adolescent), as well as the effects of sex/gender on behavioural and neurobiological endpoints of drug exposure. As a result, a thorough examination of the literature is beyond the reach of this study.
Interactions between nicotine and drugs of abuse
Nicotine is believed to be the key psychoactive ingredient of tobacco, responsible for many of the abuse-related behavioural and neurochemical effects of tobacco, such as reinforcing effects and dependency induction. Nicotine works as a complete agonist with nicotinic acetylcholine receptors (nAChRs). The nAChRs are pentameric ligand-gated receptors made up of (2–7, 9–10) and (2–4) subunits that can be arranged in a variety of ways. The majority of research on nicotine's behavioural effects has concentrated on subunits containing 42, 34, 5, 7, and 2 in mesolimbic brain regions including the ventral tegmental area (VTA) and nucleus accumbens (NAcc) (for review, see ). Nicotine's capacity to raise dopamine (DA) levels in specific brain regions has been related to a number of behavioural effects.
Animal models of substance abuse/addiction
People can combine multiple substances for a variety of reasons. One medication can, for example, enhance the positive subjective effects of another, resolve certain undesirable or adverse effects of another, or act as a stand-in for a preferred drug when it is unavailable. Preclinical studies have used a variety of animal models to test these possible interactive effects, and there is now strong empirical support for both of these theories being relevant for nicotine polysubstance use. For example, drug self-administration procedures are used to determine variables related to drug-taking behaviour, which is a key feature of polysubstance usage. It is well known that most drugs abused by humans are self-administered by animals. A medication (e.g., nicotine, cocaine, alcohol, etc.) acts as a reinforcing incentive for operant success in drug self-administration studies. Changes in the rate of acquisition of self-administration or the form and location of the self-administered drug's dose-effect curve assess changes in a drug's reinforcing effects through co-administration of, or exposure to, another drug. Drug discrimination is a preclinical model of a drug's subjective effects in humans that involves teaching animal subjects to react one way after being pretreated with a training drug and another way after being given a vehicle. The aim is to teach the subject to respond appropriately to the pretreatment interoceptive cue (i.e. discriminative stimulus) provided by the training drug. Changes in a training drug's discriminative stimulus effects are then measured by changes in the dose-response curve to the left or right, or by replacement experiments to see whether other medications reproduce the training drug's discriminative stimulus. Other procedures, such as conditioned position preference (CPP)/aversion (CPA) (thought to test rewarding/aversive properties of drugs), locomotor sensitization (a measure of neural plasticity following drug exposure), and others, may be found elsewhere.
Nicotine + alcohol interactions
The most readily available and frequently consumed drugs in the world are nicotine and alcohol. There is a high comorbidity between alcohol and nicotine use, in addition to their high rates of independent use. In fact, 80–90% of patients with alcohol use disorder (AUD) are also smokers, and cigarette use in AUDs is associated with higher rates and levels of consumption than in non-AUDs. This connection between nicotine and alcohol has also been shown in a number of preclinical studies involving various nicotine exposure and drinking conditions. Repeated nicotine injections or continuous nicotine delivery, for example, has been shown to increase animal alcohol intake. Furthermore, nicotine's effect on alcohol self-administration is often dose and time dependent, and it may be stronger after repeated nicotine exposure. In a series of studies by Lê and colleagues, these variations are clearly demonstrated. Early in therapy, repeated nicotine treatment reduced or had no effect on alcohol intake, however after several days of nicotine exposure, consumption steadily increased to levels above baseline consumption see also. If nicotine is given before alcohol, however, early in treatment, a rise in alcohol consumption can occur. For example, rats given a single pretreatment with a moderate nicotine dose (0.4 mg/kg) three hours before the initial alcohol exposure had significantly higher alcohol self-administration during the first four self-administration sessions. While it appears that nicotine can help people drink more alcohol, most studies have focused on passive, experimenter-administered nicotine injections. In comparison, a recent study examined the relationship between self-administration of intravenous (IV) nicotine and oral alcohol in rats conditioned to self-administer both drugs in regular sessions separated by 5 hours. When alcohol was available first, nicotine self-administration increased, but when alcohol was available first, nicotine self-administration decreased, implying asymmetrical associations between nicotine and alcohol on consumption. When alcohol and nicotine were available in 5-minute intervals within the same session, alcohol consumption increased dramatically relative to alcohol alone, although nicotine self-administration remained unchanged. Increases in alcohol self-administration during alternating access conditions appeared after about a week, similar to the experimenter-administered nicotine. More research is required to decide which exposure conditions encourage or discourage increased consumption of either or both substances, as well as to record the neurochemical changes that occur during nicotine exposure that contribute to increased alcohol consumption.
Nicotine can also affect variables like cessation and relapse that are linked to AUD treatment progress. Clinical studies that looked at whether one or both medications could be used in therapy yielded mixed results. Some studies have found that either quitting smoking or continuing to smoke may increase the risk of relapsing to drinking, whereas others have found that quitting smoking does not interfere with or may help in the treatment of AUD. Nicotine exposure during extinction has been shown in preclinical studies to reinstate extinguished reacting for alcohol, and this effect is stronger in subjects who have previously been exposed to nicotine (see also). Furthermore, rats conditioned to self-administer IV nicotine and oral alcohol found that co-administration of the two drugs retained self-administering levels comparable to each drug alone. When the nicotine or alcohol portion of the nicotine + alcohol condition was removed, nicotine responses faded quickly when alcohol was still present, but alcohol responses faded more slowly when nicotine was still available for self-administration. Furthermore, mice given simultaneous exposure to nicotine (P.O. in drinking water) and alcohol (daily injections) showed more somatic withdrawal symptoms that lasted longer (i.e., >72 hours) than mice given either drug alone (i.e., 48 hours). Finally, acute nicotine prevented withdrawal symptoms from alcohol cessation, and acute alcohol prevented nicotine cessation withdrawal symptoms. These results, taken together, may have significant consequences for the treatment of comorbid nicotine and alcohol use disorders, and they indicate that the order in which treatments are administered is an important factor to consider.
Several studies have been conducted to determine the behavioural basis for nicotine-induced changes in alcohol consumption. Nicotine does not substitute for the alcohol discriminative signal, and alcohol does not substitute for the nicotine cue in drug discrimination research. Nicotine, on the other hand, potentiates the discriminative stimulus effects of alcohol, especially when combined with low or threshold doses of alcohol, while alcohol attenuates nicotine's discriminative cue through see, likely through overshadowing processes, as seen in human studies. When trained as a discriminative stimulus, nicotine + alcohol combinations have also been shown to generate a discriminative cue that is distinct from the individual drugs. Drug discrimination research, taken together, can provide insight into why nicotine and alcohol are often used together in human populations. To begin with, nicotine intake (via cigarette smoking) improves the positive subjective (or discriminative stimulus) effects of low alcohol consumption 64. However, the effects of nicotine become less noticeable as more alcohol is consumed, leading to a rise in cigarette smoking. Finally, as levels of nicotine and alcohol use rise, distinct subjective effects arise.
Since nicotine did not affect alcohol-induced conditioned taste aversions 67,68 and alcohol did not affect nicotine-induced CPP 67, there is no evidence of a direct association between nicotine and alcohol in aversive or rewarding motivational steps. Nicotine and alcohol do, however, cross-reinstate extinguished CPP 69. Nicotine has been shown to reduce alcohol-induced ataxia 70,71, implying that nicotine can reduce some of the direct effects of higher alcohol doses (i.e., the descending limb of the dose-effect curve), enabling subjects to self-administer more medication. Nicotine co-administration with alcohol stimulates the production of alcohol (1 g/kg)-induced locomotor sensitization 73 and increases locomotor activity 72. Taken together, these results indicate that nicotine's ability to increase alcohol consumption is possibly due to their interaction with a wide range of behaviours.
Nicotine and alcohol tend to interact within traditional pharmacological, neurochemical, and molecular pathways, in addition to their behavioural interactions. While glutamate 74, GABA 50, and serotonin 75 may also play important roles, research has primarily focused on nAChRs in the mesolimbic DA system as key mediators of these effects. While nicotine's main focus of action is the nAChR, they also tend to be involved in many of alcohol's abuse-related effects (reviewed by 76,77). Mecamylamine, a nonselective nAChR antagonist, has been shown to minimise alcohol-induced increases in extracellular DA in the NAcc 78 and reduce alcohol consumption in rodents 79–82. Varenicline (Chantix), an FDA-approved smoking cessation aid and partial agonist of the nAChR receptor, has shown similar effects 83,84 and has been proposed as a treatment for AUD 85. Preclinical studies, on the other hand, have yielded mixed findings, with some 86–88 but not all 88–90 conditions suggesting that varenicline can minimise alcohol consumption. Varenicline also had no effect on alcohol-induced CPP, although it did affect alcohol-induced changes in locomotor activity 91.
In addiction-related brain areas 93, both nicotine and alcohol increase Fos expression (a molecular marker of neuronal activation 92) and other gene expression. The VTA has been implicated in neurobiological research as a key structure mediating the interactions between nicotine and alcohol. Alcohol injections directly into the VTA, for example, raise NAcc DA levels, which are blocked by mecamylamine 94. Furthermore, nicotine injected directly into the posterior VTA 75 or the basal forebrain, which receives feedback from the VTA 95, causes rats to seek alcohol. VTA activation also raises extracellular DA levels in the NAcc shell, and nicotine and alcohol administered directly into the VTA have an additive effect on DA levels in this area 96. Finally, human genetic studies have found shared genes linked to the development of nicotine and/or alcohol use disorders 97,98, and preclinical studies have found a connection. Rats bred for high alcohol consumption (e.g., alcohol-preferring rats) self-administer more nicotine 44,75,99 than rats bred for low alcohol consumption. Because of their modulatory function on various neurotransmitter systems (e.g., DA, GABA, glutamate, etc.) that mediate behavioural responses to both nicotine and alcohol, genes encoding for several nAChR subunits have been implicated as potential contributors (see Table 1 in 97). However, the risk of nicotine and alcohol co-use is most likely the product of a complex interaction between multiple genes 98, and further research using transgenic mouse models may shed light on the role of particular genetic alleles in various aspects of nicotine and/or alcohol co-use.
Nicotine + stimulant interactions
Clinical reports indicate an association between nicotine and a number of stimulants such as cocaine, methamphetamine (METH), and amphetamine, similar to the findings recorded of nicotine and alcohol co-use. Many stimulant abusers report a history of cigarette smoking prior to the initiation of stimulant usage 100–102, and the incidence of cigarette smoking among cocaine or METH abusers is 3–4 times that of the general population. Furthermore, cocaine-dependent smokers use more cocaine than non-smokers 100,103, and the rate of cigarette smoking rises during cocaine usage 104. Clinical trials have also discovered that using nicotine and cocaine together improves some favourable drug scores, and consumers claim that nicotine increases cocaine's reinforcing effects 105.
Medical reports and preclinical studies on the impact of nicotine on stimulant consumption have generally agreed. In rats 107,108, pretreatment with nicotine before cocaine self-administration sessions has been shown to speed up the acquisition of 106 and increase total levels of cocaine and amphetamine consumption under a progressive ratio schedule of reinforcement. Interestingly, the effects of repeated nicotine therapy take several days until cocaine self-administration rises 107, similar to the effects seen with alcohol. Nonhuman primates have also been used in studies to determine the potential associations between nicotine and cocaine when cocaine and nicotine were self-administered at the same time. In these trials, monkeys were taught to self-administer IV cocaine, then nicotine was added to the solution to compare the reinforcing effects of the mixtures to cocaine alone 109,110. Adding nicotine to cocaine solutions resulted in leftward changes in the cocaine dose-effect curves in both experiments, indicating an improvement in potency. In comparison to cocaine studies, the effects of nicotine on METH self-administration are less evident 111, but further research is required to determine what, if any, conditions of nicotine administration affect METH self-administration. One study found that acute pretreatment with a low (0.3 mg/kg) but not a heavy (1–3 mg/kg) dose of METH increased nicotine self-administration in rats 112.
Acute nicotine exposure (priming) was also tested in some experiments to see whether it could reinstate extinguished stimulant-seeking activity, but the findings were mixed 107,111,113. Hiranita and colleagues 114–116, on the other hand, have thoroughly investigated the impact of nicotine pretreatment on the reactivation of extinguished METH-seeking caused by METH priming or a METH-associated cue. The authors conclude that the nAChR system may play a role in relapse to METH-seeking since these studies consistently show that nicotine has an inhibitory effect on METH reinstatement. Nicotine administration may, interestingly, restore METH seeking after extinction, however this effect appears to be limited to rats that have previously been exposed to nicotine 111. Amphetamine self-administration was also more resistant to extinction in rats that had previously been exposed to nicotine 108.
The behavioural and neurochemical effects of nicotine and abused stimulants tend to correlate in drug prejudice research, but these effects appear to be asymmetric and drug and species based. Nicotine, in rats 117,118 and METH in rats 119 and monkeys 120, substitutes for the discriminative stimulation effects of cocaine, but in monkeys and pigeons 110,117,120, there is no replacement or partial substitution for cocaine. In rats, cocaine does not replace the nicotine discriminative stimulus 117,118.
In addition to direct changes in cocaine's reinforcing and subjective effects that tend to perpetuate or intensify drug-taking activity, cocaine users often report using nicotine-containing products following cocaine use because they "increase mental concentration" and "make them feel better physically" 121. Preclinical research also suggests that nicotine can help to mitigate some of the negative effects of long-term stimulant usage. Nicotine exposure attenuated METH-induced disturbances in cognitive function and striatal deficits, according to a recent series of findings, probably by activating 42 receptors 122–124. Nicotine and other nAChR agonists have also been shown to improve cognitive output in monkeys who have previously self-administered cocaine 125. These findings suggest that, while nicotine's positive reinforcing effects can help to sustain nicotine use, its involvement in polysubstance use may also be the product of a negative reinforcement mechanism in which nicotine counteracts some of the negative effects of repeated stimulant use. More research in this area may help to shed light on this critical problem.
Nicotine's facilitative impact on stimulant self-administration may be linked to nAChR system adaptations, at least in part. When mecamylamine was added to the cocaine solution 127, it prevented the escalation of cocaine self-administration in rats reacting to cocaine under long-access conditions, which has been stated to generate an escalation of drug intake over many sessions (see 126). When mecamylamine was withdrawn, however, cocaine use increased in the same subjects. The distribution of nAChRs is also altered in monkeys with a history of cocaine self-administration 125 and in human cocaine users 128 according to radioligand binding reports. Cross-sensitization (a neurobiological adaptive mechanism thought to play a role in the production and maintenance of addiction; 129) and other behavioural effects associated with the relationship between nicotine and stimulants have been shown. Nicotine and amphetamine 130, methylphenidate 131, cocaine 132, and METH 133 are examples of these experiences. Pre-exposed nicotine-exposed rats showed similar neurobiological improvements. Nicotine exposure, for example, increased cocaine-induced long-term potentiation and gene transcription patterns in the striatum, which may be linked to increased cocaine-induced locomotor sensitization and CPP 14. Nicotine's behavioural and neurochemical effects, on the other hand, were not improved by cocaine use. Epidemiological evidence supports these laboratory results, indicating that most cocaine users begin using the drug after a history of tobacco smoking and while still smoking cigarettes 14. These results have led some to label nicotine a "gateway drug" 134, but this label has yet to be thoroughly tested, and some concerns remain about its applicability to human drug users.
The mesolimbic DA system, especially the NAcc and other striatal regions 31,135, has been the focus of research into the neurochemical basis for nicotine + stimulant interactions. In rats conditioned to self-administer nicotine or cocaine, both nicotine and cocaine produce overlapping patterns of activation in mesolimbic DA pathways 136. Nicotine, like cocaine and METH, raises extracellular DA levels in the NAcc 32,137, according to microdialysis reports. Importantly, the effect of nicotine and cocaine on extracellular DA is often referred to as additive 138–140, which corresponds to the degree of improvement in most behavioural steps. However, as compared to stimulant substances including cocaine and METH (>600 percent), the maximal increases in extracellular DA levels caused by nicotine are relatively minor (typically about 100–200 percent increase from basal values). Furthermore, positron emission tomography (PET) experiments in rhesus monkeys revealed that nicotine does not release enough DA to displace 11Craclopride from DA receptors, even at doses that increase cardiovascular responses. METH, on the other hand, generates a stable, time-dependent displacement 141, see also 142. Nicotine-induced increases in extracellular DA may be sufficient to produce stimulant-like effects, even if they are small. In rats trained to discriminate cocaine 143 or METH 144 from saline, a low dose of the training drug that produced > ~125% of baseline DA in microdialysis studies was enough to produce stimulant-like discriminative stimulus effects suggesting a role for nicotine-induced DA release in its abuse-related effects and interactions with other drugs of abuse.
Nicotine + opioid interactions
According to clinical studies, 83–95 percent of methadone maintenance patients smoke cigarettes 145,146, and there has also been a connection between smoking status and prescription opioid abuse 147. Nicotine and opiates (e.g., heroin, morphine, methadone) combine in laboratory-based human self-administration experiments to increase overall drug intake. For instance, the administration of heroin 8 or methadone 148 increases smoking, while methadone also increases subjective ratings of smoking satisfaction 148. The opioid antagonist naltrexone reduces the amount of cigarettes smoked as well as self-reported smoking satisfaction 149. Finally, laboratory tests have shown that after nicotine exposure, subjects consume more methadone 150,151. These findings could explain the high smoking rates among methadone-dependent patients 152.
Although nicotine self-administration in rats does not appear to be dependent on mu opioid receptor activity, antagonists such as naloxone or DAMGO do not alter nicotine intake 153,154, preclinical studies have not specifically assessed the effects of nicotine on opiate self-administration (or vice versa). Other opioid reward measures, on the other hand, have been investigated. In rats, intracranial self-stimulation studies (ICSS) show that morphine pretreatment lowers the ICSS threshold for nicotine, indicating improved efficacy and potency 155. Nicotine and opiates can cause cross-tolerance 156 or cross-sensitization 157, which means that, depending on the schedule and period of administration, exposure to one drug increases or decreases the reaction to the other in CPP procedures. Furthermore, acute nicotine administration into the VTA 158, hippocampus 159, or basolateral amygdala 160 enhances morphine-induced CPP, while mecamylamine inhibits it. Morphine also restores the nicotine-induced CPP 161 that had been lost. Interestingly, systemic or central (ICV) injections of “ultra-low doses” of nicotine (0.0012–0.01 mg/kg, intraperitoneal; 0.03–0.5 ng/mouse, ICV) have been shown to minimise the expression of morphine-induced CPP 162. Although there is no clear explanation for these results, they may be related to memory disturbances, since these same nicotine doses have been shown to affect morphine state-dependent learning 163.
There is no variation in the discriminative stimulation impact of nicotine and opioids, according to drug discrimination reports. Morphine, for example, does not replace nicotine's discriminative stimulus effects 164, and nicotine does not replace morphine's discriminative stimulus 165. Furthermore, naloxone and naltrexone, both mu-opioid antagonists, had no effect on nicotine's discriminative cue 165,166, and mecamylamine had no effect on morphine's discriminative cue 166.
The apparent overlapping mechanisms in the development of tolerance and/or abstinence from prolonged treatment with nicotine or opiates 167 has gotten a lot of attention. In morphine-treated mice, repeated nicotine treatment has been shown to reduce the production of morphine tolerance as measured by less opioid antagonist-precipitated withdrawal signs including withdrawal jumping 168 or CPA 169,170. Acute nicotine also reduces opioid withdrawal caused by antagonists 171,172. While nicotine withdrawal symptoms tend to be milder than opiate withdrawal symptoms 173, several studies have discovered that endogenous opioids are involved in nicotine tolerance and withdrawal. Opioid antagonists, for example, inhibit nicotine tolerance 174, precipitate nicotine withdrawal in mice 174,175, and obstruct nicotine-induced nicotine withdrawal alleviation 176. Mu opioid receptor agonists 175, including nicotine, reduce withdrawal symptoms.
Nicotine-induced improvements in the reinforcing effects of opioids, including nicotine's interactions with other medications, are possibly attributable to changes in the mesolimbic DA mechanism, as morphine-induced DA release in the caudate putamen and NAcc is greatly augmented by chronic nicotine administration 157. Nicotine and opiates have interactions that go beyond the drugs' reinforcing effects to other essential effects like antinociception, but they aren't mutually exclusive because people with chronic non-malignant pain often use opioids and nicotine to control their pain (i.e., negative reinforcement; 177). There's evidence that nicotine releases endogenous opioids 178,179 and that nAChRs play a role in opioid-induced antinociception 180, possibly by potentiating opioid-induced antinociception through nAChR-induced endogenous opioid peptides 181. To recognise the importance of this relationship to nicotine + opioid co-use, preclinical pain models must be used to identify the essence of nicotine's reinforcing impact.
Nicotine + THC interactions
Nicotine and THC usage have a clear link in humans, and cannabis users say that nicotine enhances the effects of THC, the psychoactive component in marijuana 182. In addition to these epidemiological observations, a double-blind cross-over experiment using transdermal nicotine (21 mg) significantly altered the physiological and subjective effects of smoked marijuana (1.99 or 3.51 percent THC content). Participants reported significantly higher increases in heart rate and subjective experiences of being "stimulated," as well as the fact that THC's effects lasted longer 183.
In terms of rewarding/reinforcing effects, preclinical studies have shown a similar relationship between THC or other cannabinoid (CB) receptor agonists and nicotine. In nicotine-induced CPP studies, the CB receptor antagonist rimonibant (SR141617A) blocked the acquisition of nicotine CPP 184 as well as nicotine priming-induced reinstatement of extinguished nicotine CPP 69. In CB1 receptor knockout mice 185, nicotine fails to condition CPP, suggesting that the endocannabinoid system is involved in nicotine reward. In a self-administration study 186, rats were given escalating THC doses for three days before learning to self-administer nicotine seven days later. THC-exposed rats had a higher rate of nicotine self-administration (94%) than vehicle-exposed rats (65%), and THC-exposed rats had self-administered more nicotine after acquisition. WIN55,212–2 and other CB1/2 agonists have been shown to improve nicotine self-administration and restore nicotine-seeking activity. Number 187. Rimonabant was found to reduce nicotine self-administration in rats 188. Since CB receptor agonists and antagonists have no impact on nicotine's discriminative stimulus effects, these increases in nicotine self-administration are most likely due to reinforcing effects 188,189. (see 190). As of this writing, no studies seem to have looked into the effects of nicotine on THC self-administration. Nonetheless, evidence indicates that activation of CB receptors can affect various aspects of nicotine's rewarding and reinforcing effects.
Nicotine can help THC's other pharmacological and behavioural effects. Nicotine, for example, assisted the hypothermic, antinociceptive, and hypolocomotive impact of THC administration in mice 191. Furthermore, combining nicotine and THC decreased THC-induced tolerance while increasing the expression of the THC withdrawal-precipitated CB receptor antagonist 191. THC reduced the frequency of nAChR antagonist-precipitated nicotine withdrawal (wet dog shakes, paw tremors, scratching) and reversed CPA associated with antagonist-precipitated nicotine withdrawal 192. Finally, studies on rodent drug discrimination have discovered that nicotine improves low-dose THC discrimination 193.
Several lines of evidence point to a connection between the endocannabinoid and nicotinic acetylcholine systems, which may clarify the pharmacological and behavioural interactions between THC and nicotine. 194 conducted a review of the evidence for the endocannabinoid system's role in nicotine addiction. It included studies in molecular biology, pharmacology, and behaviour. Although interactions between nicotine and alcohol or nicotine and abused stimulants tend to be linked to increases in extracellular DA in the NAcc (see above), the relationship between nicotine and THC is likely to be more complex, with studies showing that CB receptor antagonists minimise nicotine-induced mesolimbic DA release but have no impact on NAcc DA release 188,195. More research is needed to fully comprehend the dynamic interactions between nicotine and THC on the DA system.
Cocaine Toxicity:
Cocaine remains one of the most common triggers of drug-related emergency department (ED) visits in the United States, despite being dominated by opioids in recent years. Cocaine toxicity can affect virtually any organ system, but the majority of patients present with cardiovascular symptoms.
Body packers—people who swallow cocaine bags in order to smuggle the drug from one country to another—may experience acute toxicity if any of the bags burst. See the illustration below.
Two or more isoprene molecules are connected together in the biosynthesis of terpenes. Provided that the head and tail of the molecule are mainly involved in the linking, there are three possibilities for linking between two isoprene molecules:
4.4.1 Linkage
One isoprene molecule's head may connect to the head of another isoprene molecule.

Figure: This link is called a head-to-head or 1-1 link.
4.4.2 Linkage
One isoprene molecule's head may connect to the tail of another isoprene molecule.

Figure: This link is called a head-to-tail or 1-4 link.
4.4.3 Linkage
One isoprene molecule's tail may connect to the tail of another isoprene molecule.

Figure: This link is called a tail-to-tail or 4-4 link.
Crosslinks are found in cyclic terpenes that are not 1-1, 1-4, or 4-4.
eg. 1:
Figure: Myrcene
eg. 2:
Figure: Limonene
eg. 3:
Figure: Retinol
An irregular terpene is one that does not follow the isoprene law. as an example:
Figure: β-Carotene
See also isoprene unit
4.4.4 Natural occurrence:
Because of their high reactivity, most metals are found in nature in a combined state. Minerals are metals found in a mixed state in the earth's crust. Ores are minerals that have a relatively high metal content and can be processed in a simple and cost-effective manner.
Gold and platinum are the only metals that can be found in their natural state. They are commonly embedded in the earth's crust with earthly (or) rocky impurities like sand and clay.
Metal abundance varies in the earth's crust.
By weight, the relative abundance of certain metals in the earth's crust is:
Metal | Relative Abundance |
Al | 8.23 |
Aluminium and iron are more plentiful than other metals. By weight, aluminium accounts for around 8% of the earth's crust, while iron accounts for 5.8%.
As a result, aluminium is the most abundant metal in the earth's crust, and iron is the second most abundant metal by weight. In addition, by weight of the earth's crust, aluminium is the third most abundant element, and iron is the fourth most abundant element.
The main ores of certain essential metals, as well as their chemical composition:
Metal | Ores | Composition |
Aluminium (Al) | Bauxite, | AlOx(OH)3-2x [Where 0<x<1] |
Iron (Fe) | Haematite | Fe2O3 |
Copper (Cu) | Copper pyrites | CuFeS2 |
Zinc (Zn) | Zinc blende or Sphalerite | ZnS |
Aluminium and iron are commonly used in industrialised countries. Aluminium is a major component of silicates like mica and clay that occur naturally. Aluminium is mostly derived from bauxite, which is the primary ore.
Aluminium oxides, as well as traces of transition metals, are found in valuable gem stones like rubies and sapphires. Rubies are crystalline types of aluminium oxide that are reddish-brown in colour due to chromium impurities. Due to the presence of impurities such as nickel, cobalt, and titanium, sapphires come in a variety of colours.
Haematite and magnetite are the most common oxide ores from which iron is extracted. The iron extracted from these ores is primarily used to make steel and iron alloys. Iron is also a necessary component of biological systems.
Copper and zinc are two other metals that are widely used.
4.4.5 Elements of modern periodic table:
Five elements make up the group 16 elements of the current periodic table: oxygen, sulphur, selenium, tellurium, and polonium. Since several elements can be extracted from sulphide or oxide ores, these elements are also known as chalcogens or ore-forming elements.
On the surface of the earth, oxygen is available. After hydrogen, helium, and neon, oxygen was believed to be the fourth most abundant element in the universe, based on the proportions of various forms of atoms present in the universe. It makes up 89 percent of water, 46 percent of the earth's crust, and 20% of the atmosphere.
Period | Element | Symbol | Atomic Number | Electronic Configuration |
2 | Oxygen | O | 8 | [He] 2s2 2p4 |
3 | Sulphur | S | 16 | [Ne] 3s2 3p4 |
4 | Selenium | Se | 34 | [Ar] 3d10 4s2 4p4 |
5 | Tellurium | Te | 52 | [Kr] 4d10 5s2 5p4 |
6 | Polonium | Po | 84 | [Xe]4f14 5d10 6s2 6p4 |
Oxygen:
Oxygen is represented by the chemical symbol O. It is a colourless, odourless gas that is converted into carbon dioxide during the human respiration process. The molecule of oxygen is a diatomic molecule (O2). In traces, oxygen can also be present as a triatomic molecule (O3), which is known as ozone. Many elements mix easily with oxygen. The evolution of heat energy occurs during the combination of certain components, and this process is known as combustion.
Sulphur:
Sulphur is represented by the letter S. It is a non-metal that comes in ninth position in terms of celestial abundance. Sulphur atoms make up about one in every 20,000-30,000 atoms. Sulphur can be present in both the combined and free states. Seawater contains around 0.09 percent sulphur in the form of sulphates. A considerable volume of sulphur is contained in underground deposits of pure sulphur present in dome-like structures, and the meteorite contains 12 percent sulphur. The activity of anaerobic bacteria on sulphate minerals such as gypsum produces sulphur.
Selenium:
Selenium is more scarce than either oxygen or sulphur. In a few minerals, it can be found both free and mixed with heavy metals (such as iron, silver, or mercury). Under normal conditions, the grey metallic form of selenium is the most stable form of the element.
Tellurium:
Tellurium is a chemical element with the atomic number 52 that has properties that are similar to both metals and non-metals. It's one of the rarest stable elements on the planet's surface. It's commonly found in free form and in compounds with other elements including copper, lead, silver, and gold.
Polonium:
Among the group 16 elements, it is the rarest. It's a radioactive substance. Polonium is also used for alpha radiation in research applications.
4.5.1 Structure of Citral:
2D
3D
Crystal Structure:
4.5.2 Synthesis:
Citral is a substance that can be used to produce vitamin A. This industrial synthesis is made up of two [3,3]-sigmatropic rearrangements that are carried out in order.
The first is a Claisen rearrangement, which occurs when an oxygen is present in a chair-like transition state. More information about the Claisen reorganisation can be found here. The Cope-rearrangement is the second part of the single-step reaction, in which only carbons are involved in the chair-like transition state. When the two starting materials are heated together, these two reactions occur in succession, and the reaction is fueled by the formation of a conjugated carbonyl group in the substance.
4.5.3 Structure of Neral:
2D
3D
4.5.4 Structure of a-terpionel
2D
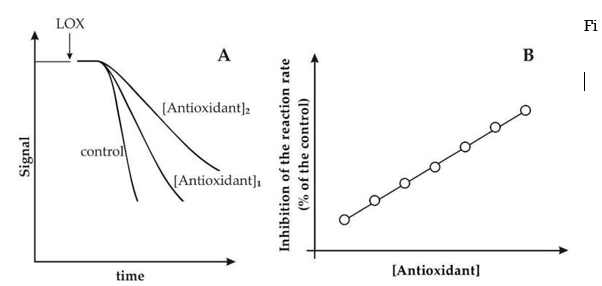
3D
4.5.5 Synthesis of a-terpineol:
Terpineol is widely made from the more readily available alpha-pinene, despite the fact that it is naturally occurring. A different path begins with limonene:
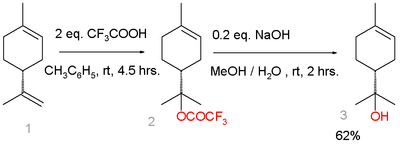
● Terpineol synthesis from limonene
In a Markovnikov reaction, limonene reacts with trifluoroacetic acid to form a trifluoroacetate intermediate, which is quickly hydrolyzed with sodium hydroxide to yield -terpineol with a 7 percent selectivity. -terpineol in a combination of the cis and trans isomers, as well as 4-terpineol, are side-products.
Geranyl pyrophosphate, which releases pyrophosphate to give the terpinyl cation, is used in the biosynthesis of -terpineol. Many terpenes and terpenoids are formed from this carbocation. Terpineol is generated when it is hydrolyzed.
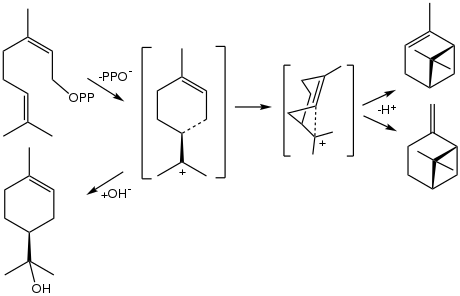
Geranyl pyrophosphate biosynthesis to the terpenes -pinene and -pinene (right) and -terpineol (left) (bottom left).
References:
1. Graham Solomons T. W., Fryhle, Craig B., Snyder Scott A, Organic Chemistry, Wiley
Student Ed, 11th Edition (2013)
2. Jonathan Clayden, Nick Greeves, Stuart Warren, Organic Chemistry, 2nd Edition, Oxford
Publisher, 2014.
3. Dhawan, S.N., Pradeep’s Organic Chemistry, (Vol. I and II), Pradeep Publications




